

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们导读
核糖体通过小亚基的解码中心和大亚基的肽酰转移酶中心调控基因表达。在酿酒酵母中,影响18S或25S rRNA的突变通过非功能性rRNA降解(NRD)途径下调,分为18S NRD和25S NRD。尽管这已在酿酒酵母中发现,但NRD的机制在哺乳动物中尚不清楚。
细胞应对压力时,会通过eIF2α磷酸化抑制蛋白质合成,激活eIF2α激酶,启动整合应激反应(ISR),减少翻译起始,并上调ATF4。GCN2可通过碰撞核糖体激活,但其机制仍未明了。
核糖体停滞时,RQC途径激活,停滞核糖体与后续核糖体碰撞,E3泛素连接酶ZNF598识别并泛素化核糖体蛋白,促进mRNA切割和核糖体回收。解决核糖体的碰撞有助于避免ISR和免疫反应。
核糖体蛋白uS3和uS5的泛素化与细胞应激相关,E3泛素连接酶RNF10通过泛素化靶向清除停滞核糖体。酵母中RNF10的同源蛋白Mag2可识别停滞的80S核糖体,提示18S NRD可能存在于哺乳动物中,RNF10也可能参与其中。
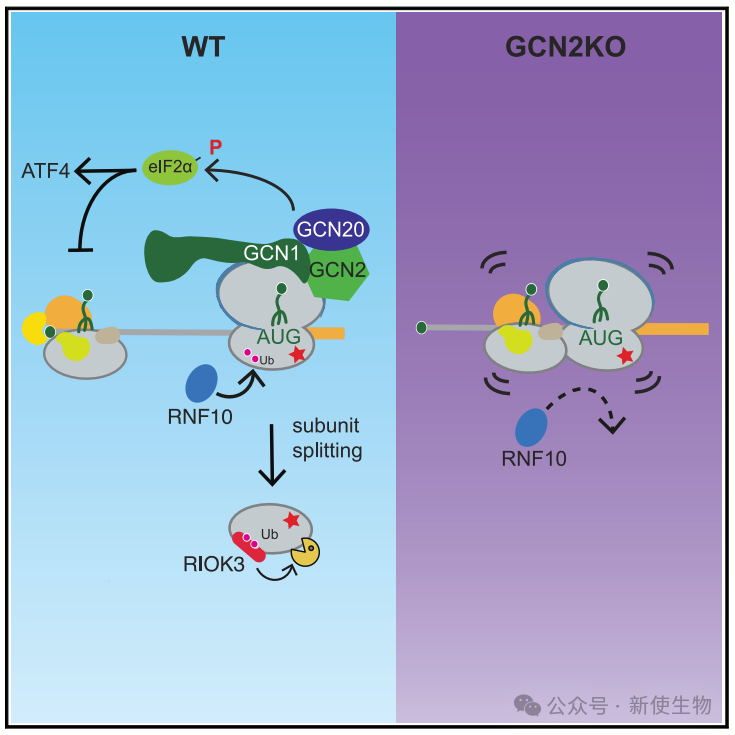
2025年2月12日,美国国立卫生研究院Colin Chih-Chien Wu团队在Molecular Cell上发表了一篇题为“The integrated stress response regulates 18S nonfunctional rRNA decay in mammals”的论文,发现哺乳动物18S NRD通过整合应激反应由GCN2启动。非功能性18S rRNA导致其在起始位点处翻译停滞,揭示了GCN2-RNF10-RIOK3轴在翻译起始阶段监控核糖体功能性的动态反馈机制。

文章索引
【发表日期】2025年2月12日
【IF】15.58
研究结果
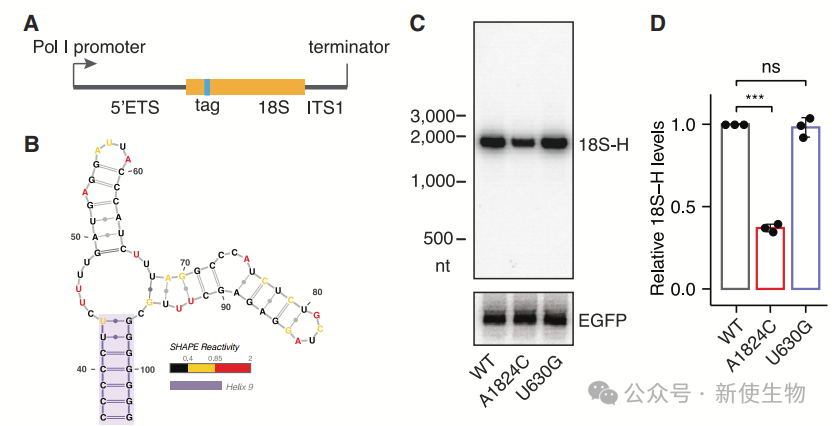
一. 研究人类18S NRD的正交rRNA表达系统
作者开发了正交人类18S rRNA表达系统,发现A1824C突变使18S rRNA在哺乳动物细胞中下调,并加速降解。
实验结果表明,哺乳动物细胞中存在18S NRD,为研究这一过程提供了有效模型。
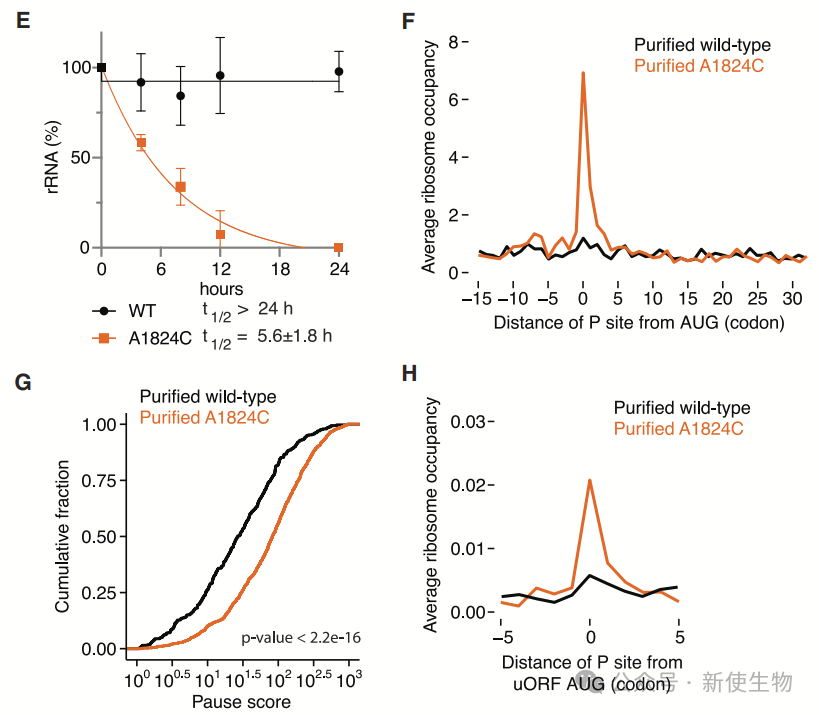
二. 解码无功能的核糖体主要停滞在翻译起始位点
Polysome profiling多聚核糖体分析显示18S:A1824C rRNA可组装成成熟80S核糖体,但因解码失能,导致核糖体主要停滞于转录组的起始密码子处。
Ribo-seq核糖体印迹分析显示核糖体富集于主ORF和uORF起始位点。
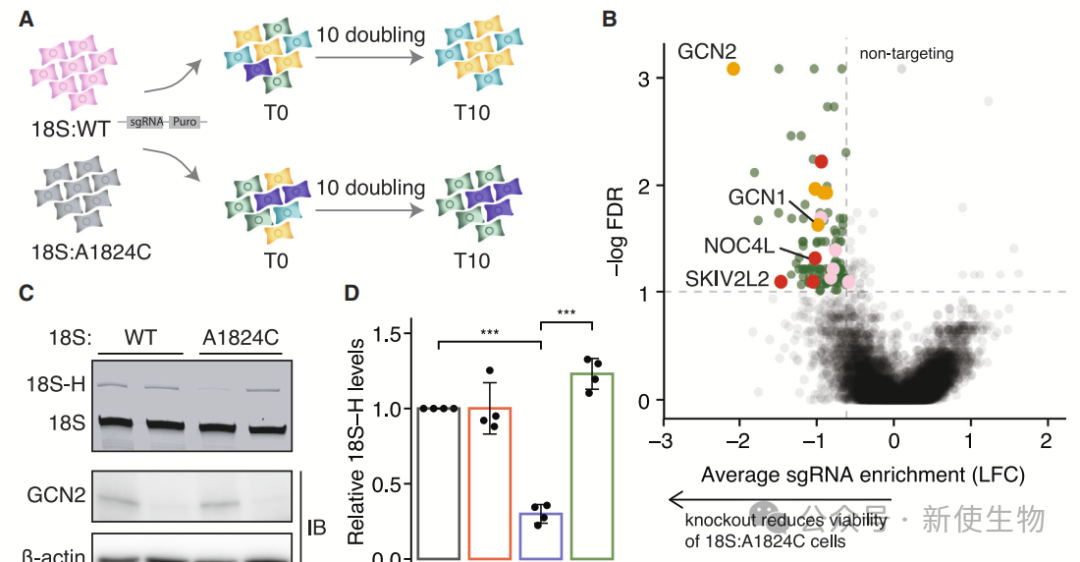
三. 基于适应性的CRISPR筛选鉴定GCN2为NRD组成部分
在正交18S rRNA表达系统中,表达18S:A1824C rRNA的细胞表现出生长缺陷。
通过CRISPR-Cas9筛选,作者发现GCN2作为关键基因,敲除GCN2会加剧18S:A1824C细胞的生长缺陷,表明GCN2参与响应非功能性18S rRNA。
四. GCN2及其效应因子在18S NRD中必不可少
GCN2缺失或GCN1敲低导致18S:A1824C rRNA积累,表明GCN2对18S rRNA降解至关重要。
核糖体P-stalk的缺失也抑制18S NRD,进一步支持GCN2与P-stalk的相互作用。
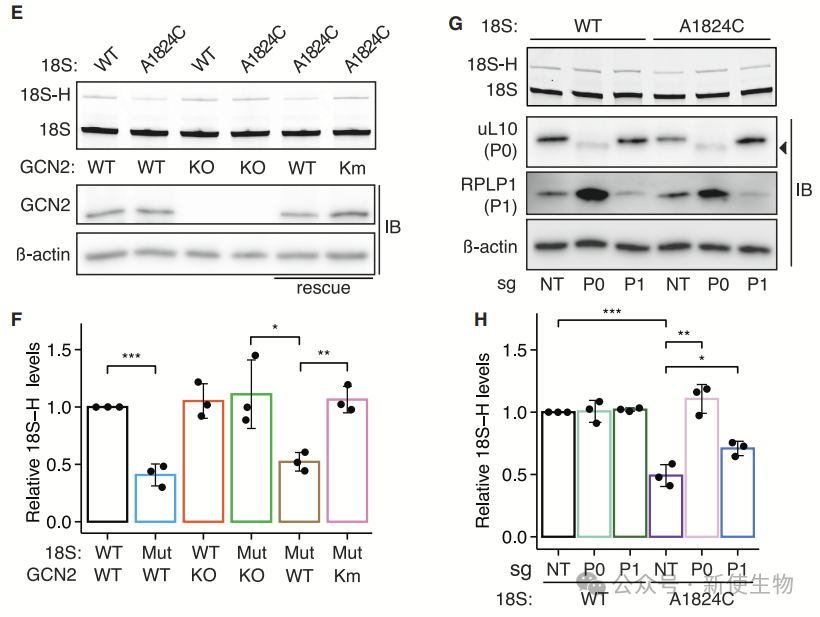
五. 解码无功能核糖体触发GCN2依赖的eIF2α磷酸化,而不引发核糖体碰撞
解码无功能的核糖体通过空缺的A位点触发GCN2依赖的eIF2α磷酸化,激活胁迫反应(ISR),但不引起核糖体碰撞。
GCN1优先结合这些解码无功能的核糖体,增强GCN2介导的翻译抑制。

六. GCN2通过精细调节翻译起始确保18S NRD
研究表明,解码无功能的核糖体通过GCN2依赖的eIF2⍺磷酸化减少翻译起始,促进18S NRD。
实验中,通过表达eIF2⍺S51D磷酸化突变体,发现中等水平的eIF2⍺S51D促进18S NRD,而非磷酸化的eIF2⍺S51A则无效。这表明翻译起始的最佳调节对于非功能性18S rRNA的降解至关重要。

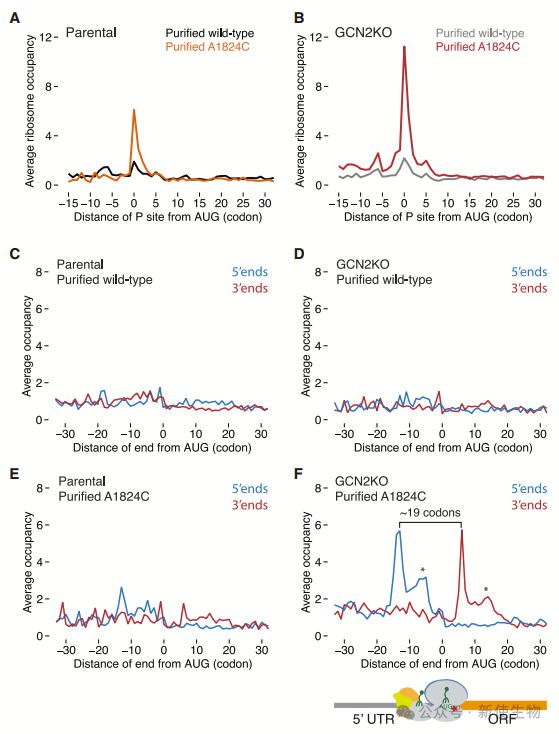
七. GCN2丧失加剧起始密码子停顿,导致43S-80S碰撞
Ribo-seq显示GCN2KO细胞中,解码无功能的核糖体在起始密码子处富集,且出现43S-80S核糖体碰撞。通过GCN2调控翻译起始,减缓这些碰撞,从而促进18S NRD。
这表明,GCN2通过抑制翻译起始,减轻碰撞,维持翻译平衡。
八. 无功能核糖体的泛素化依赖于GCN2-RNF10轴
研究发现,RNF10通过泛素化uS3促进解码无功能核糖体的降解。GCN2KO减少了uS3泛素化,表明GCN2与RNF10在18S NRD中存在遗传相互作用。
CRISPR筛选进一步确认RNF10与GCN2之间的正向遗传相互作用,支持GCN2通过RNF10介导18S NRD。

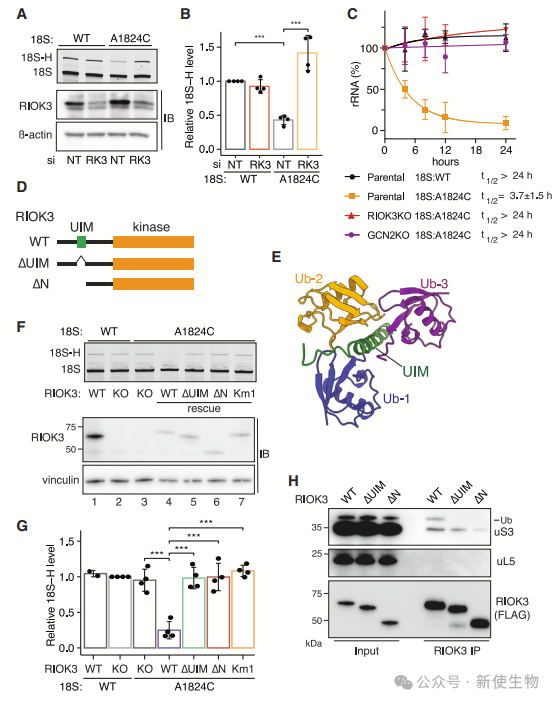
九. RIOK3的UIM和激酶活性在18S NRD中必不可少
RIOK3对18S NRD至关重要,其UIM和激酶活性必需,缺失这两者的突变体无法恢复18S NRD。
RIOK3通过UIM结合泛素化的40S亚基,激酶活性则对NRD起关键作用。
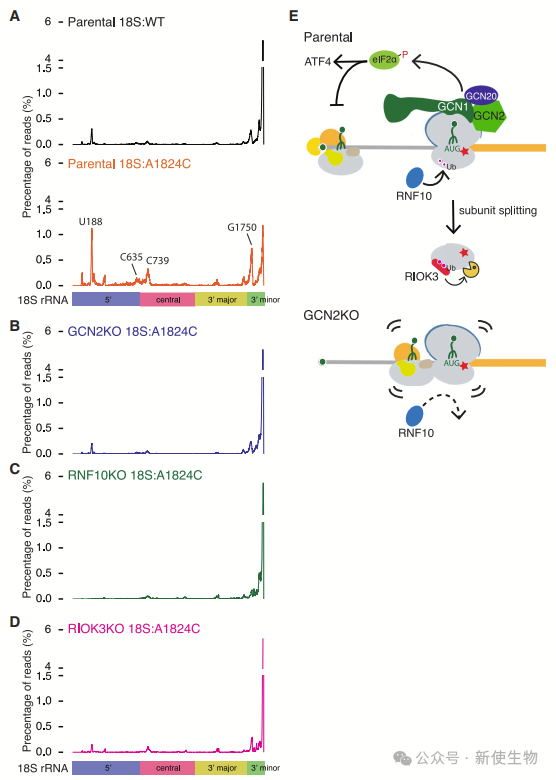
十. 18S NRD中间体的探究
作者发现18S rRNA衰变中间体主要位于5'、中心和3'次要结构域,GCN2KO、RNF10KO和RIOK3KO背景下这些中间体的形成被抑制。
RIOK3优先结合这些衰变中间体,特别是在40S亚基的5'、中心和3'次要结构域。
总结
18S非功能性rRNA衰变在哺乳动物中通过GCN2启动,激活ISR反应,限制翻译起始并减少核糖体碰撞。RNF10通过泛素化促进18S rRNA衰变,RIOK3结合泛素化的40S亚基加速其降解。此过程形成一个动态反馈机制,通过GCN2-RNF10-RIOK3轴监控核糖体功能。
超高的准确性为研究非经典的开放阅读框(ORFs)提供极大便利,提高微肽(肿瘤新生抗原)的挖掘效率。
简便快速的操作让翻译组分析不再设有门槛,每位科研学者都能轻松上手。
另外新使生物提供多物种多聚核糖体分析(Polysome profiling),了解更多翻译组技术信息可登录 www.neoribo.com

点击图片查看
点击图片查看