

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
营养剥夺会触发细胞的代谢应激通路并启动适应性程序以维持细胞存活。在长期应激下,细胞可进入一种可逆的非增殖休眠状态,其特征是代谢活性低下,从而在长期缺乏营养或水分的情况下存活。
这种休眠现象广泛存在于真菌、原生动物、植物、动物甚至癌细胞等多种真核系统中。在指数生长期,酵母将高达50%的细胞能量用于蛋白质合成,而这一过程在诱导细胞休眠的不利代谢条件下会被迅速下调。
核糖体关闭是下调蛋白质合成不可或缺的环节,该过程由一组被称为冬眠因子的特化蛋白质介导,它们不仅阻断翻译还能保护核糖体免受核酸酶降解。当环境恢复有利条件时,这些冬眠因子会解离或被取代,从而使翻译得以恢复并重新启动蛋白质合成。
2026年5月13日,弗吉尼亚大学Ahmad Jomaa与欧洲分子生物学实验室Simone Mattei团队合作,在Nature上发表了题为“SNOR promotes translation restart after dormancy” 的研究论文。本研究在粟酒裂殖酵母中鉴定出一种含有SBDS结构域的核糖体结合因子SNOR,它在葡萄糖消耗诱导的休眠期间与核糖体肽基转移酶中心结合。

文章索引
【标题】SNOR promotes translation restart after dormancy
【发表期刊】Nature
【发表日期】2026年5月13日
【作者及团队】弗吉尼亚大学Ahmad Jomaa与欧洲分子生物学实验室Simone Mattei团队
【IF】48.5
研究结果
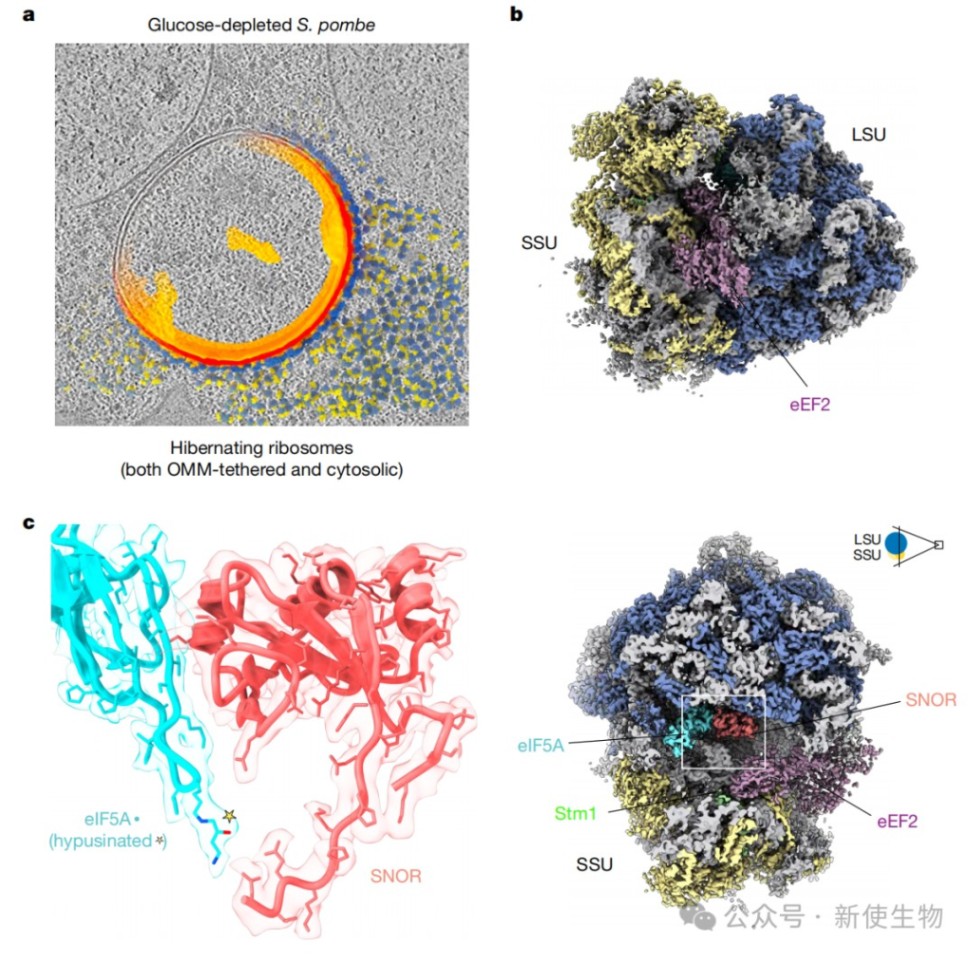
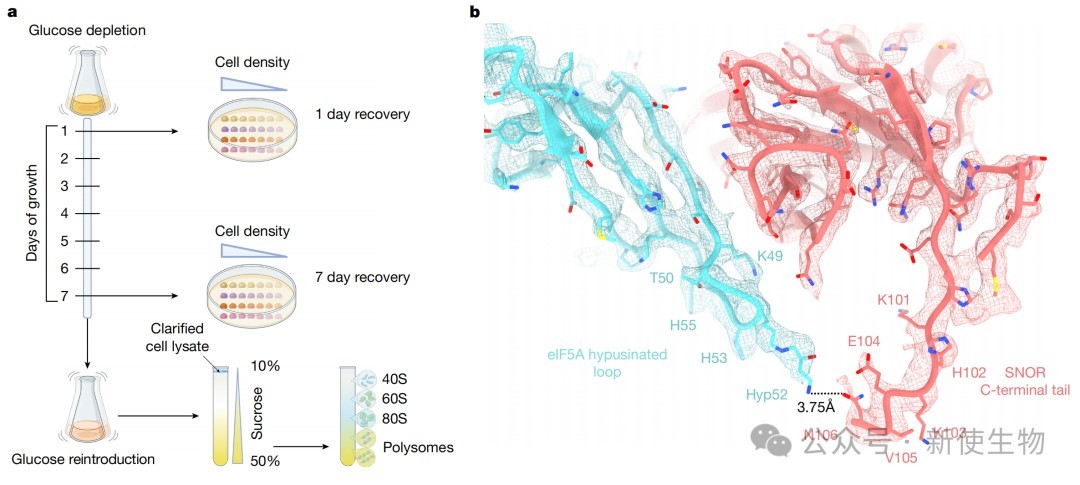
一、原位冷冻电镜断层扫描技术在休眠核糖体上鉴定出SNOR
通过对葡萄糖耗尽的裂殖酵母细胞进行高分辨率原位冷冻电镜断层扫描,研究人员在处于休眠状态的核糖体上解析出一个新的密度。
结合结构比对和质谱分析,最终确定这是一个之前功能未知的含SBDS结构域的蛋白(Rtc3),并将其命名为SNOR。
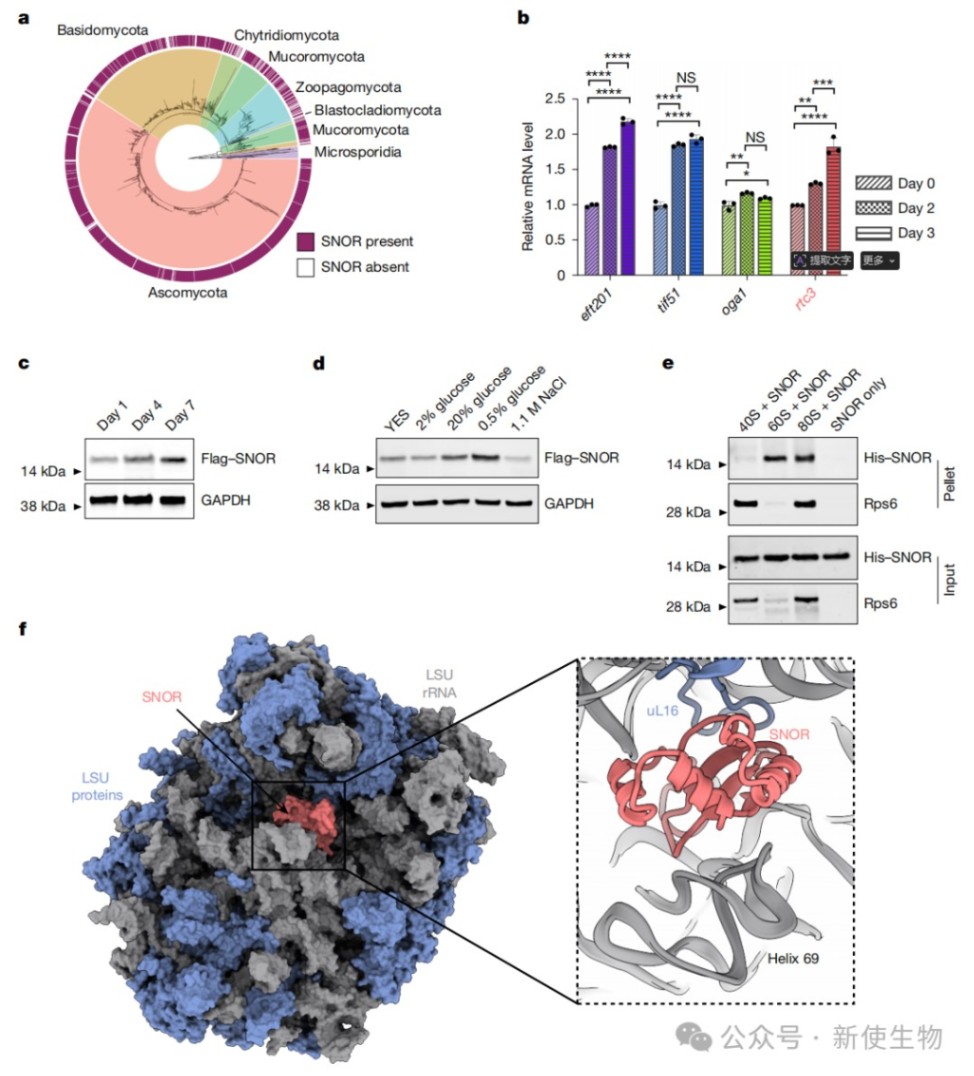
二、SNOR在葡萄糖胁迫下表达上调并与核糖体结合
通过生物信息学、RT-qPCR和免疫印迹实验,研究发现SNOR在真菌中高度保守,其mRNA和蛋白质水平均在葡萄糖限制的碳源胁迫下特异性上调。
体外结合实验证实,SNOR能够直接与核糖体大亚基(60S)和完整的80S核糖体结合。
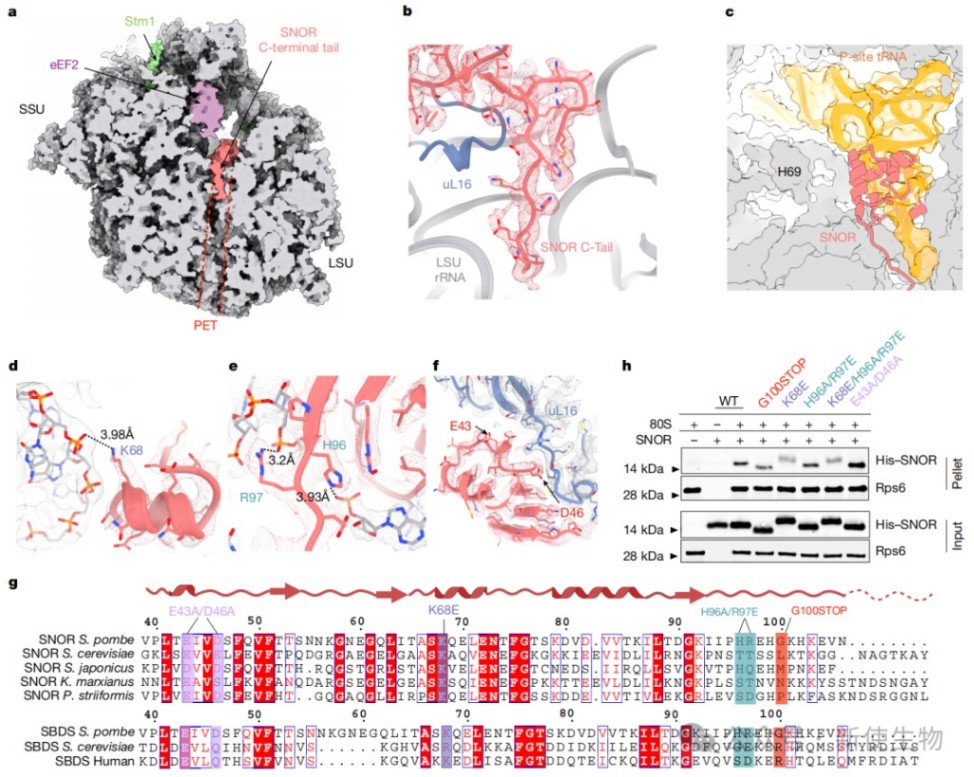
三、结构和突变分析揭示了SNOR与核糖体结合的关键位点
通过体外重构和单颗粒冷冻电镜技术,研究解析了SNOR与60S核糖体亚基复合物的精细结构,发现SNOR结合于肽基转移酶中心(PTC),其C端尾部插入多肽输出通道(PET)。
进一步的定点突变实验表明,SNOR通过其表面的特定带电残基(如K68)与核糖体RNA相互作用,这是其稳定结合核糖体的关键。
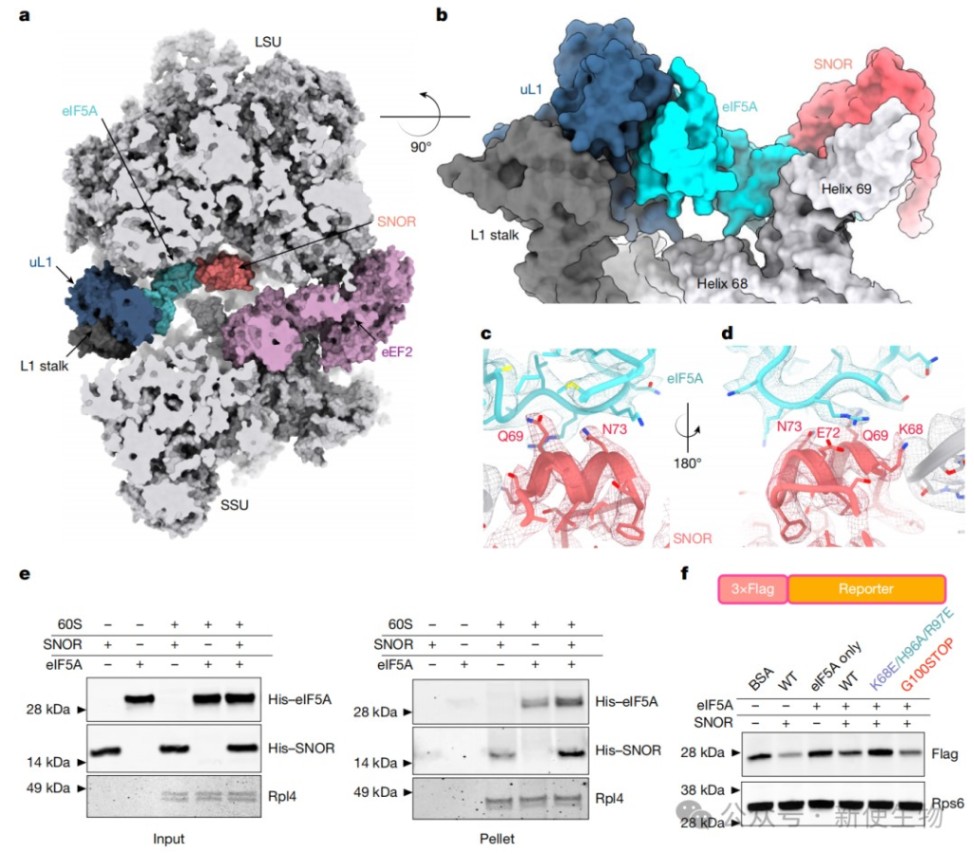
四、SNOR、eIF5A和uL1形成三方复合物以稳定核糖体
结构分析显示,SNOR与翻译因子eIF5A及核糖体蛋白uL1形成一个三方互作界面,共同稳定L1茎环等核糖体活性位点的构象。
体外结合实验证实,SNOR和eIF5A存在协同结合效应,能够增强彼此在核糖体上的稳定性。
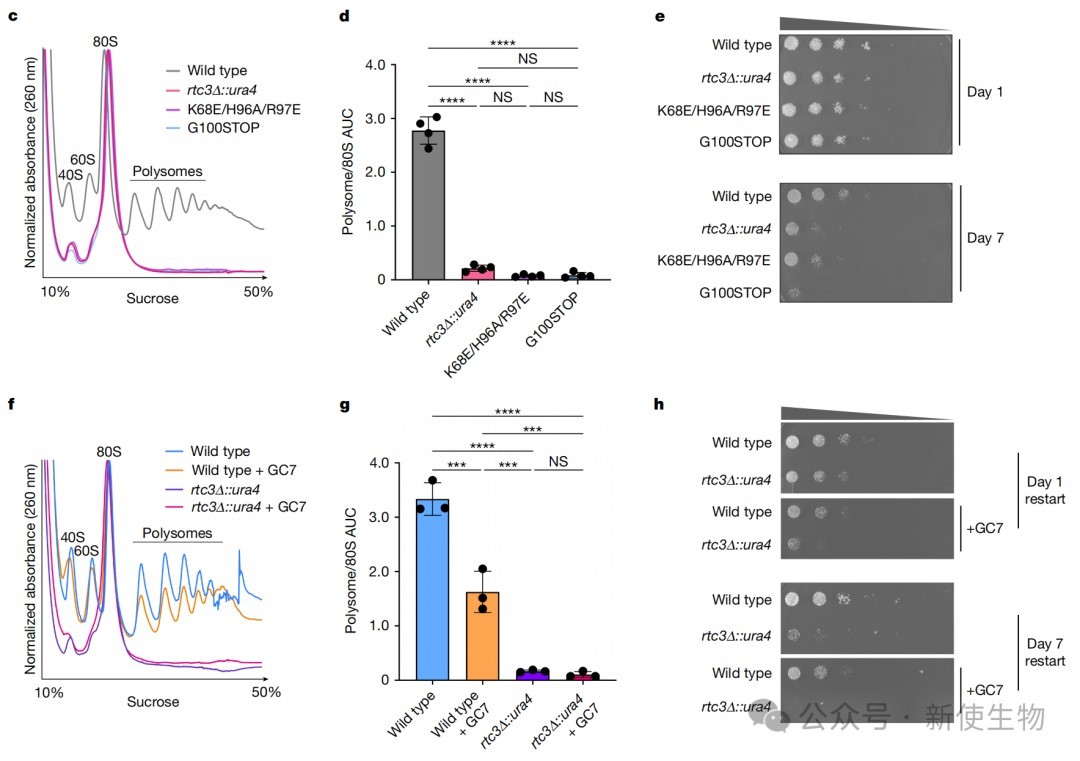
五、SNOR与eIF5A协同作用促进翻译重启和细胞存活
通过Polysome profiling多聚核糖体分析和细胞存活实验,研究发现敲除SNOR或突变其关键结合位点及C端尾巴,会导致细胞在经历长期葡萄糖饥饿后无法有效恢复蛋白质合成,并显著降低存活率。
同时,抑制eIF5A的羟腐胺赖氨酸修饰也会产生类似缺陷,证明SNOR与修饰后的eIF5A协同作用,共同促进翻译重启。
总结
本研究鉴定了一个新的胁迫响应因子SNOR,并阐明了其在细胞休眠后翻译重启过程中的关键作用。SNOR并非简单地抑制翻译,而是作为一个核糖体监视和重启的授权模块,与eIF5A协同作用,将碳源限制信号与核糖体活性位点的状态偶联起来,确保细胞在逆境解除后能迅速恢复生机,这为理解真核生物翻译调控的动态性和复杂性提供了新的见解。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看