

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
癌症免疫疗法通过增强免疫系统识别和清除恶性细胞的能力,彻底改变了肿瘤学治疗格局。尽管癌症疫苗、过继性细胞转移和免疫检查点抑制剂等策略取得了显著临床成功,但其在不同肿瘤类型中的疗效仍存在不一致性。
低突变负荷或免疫抑制性微环境的肿瘤常表现出新抗原呈递不足、T细胞浸润不良及治疗耐药等问题。此外,免疫相关不良事件的发生、高昂的治疗成本以及患者反应的异质性,均对现有疗法提出了严峻挑战。
临床迫切需要开发具有更高精准度、安全性和适应性的下一代免疫疗法。合成生物学通过构建可编程的活体治疗药物,为解决上述临床痛点提供了全新的设计范式。
近日,华东师范大学叶海峰和管宁子团队在Cell Chemical Biology上发表了题为“Programmable living therapeutics for cancer immunotherapy”的综述文章。该综述系统性地总结了合成生物学在三种主要活体治疗药物(工程化CAR-T细胞、溶瘤细菌和溶瘤病毒)中的最新突破,揭示了如何通过基因回路赋予这些系统体内感知、计算和执行的能力。

综述整理
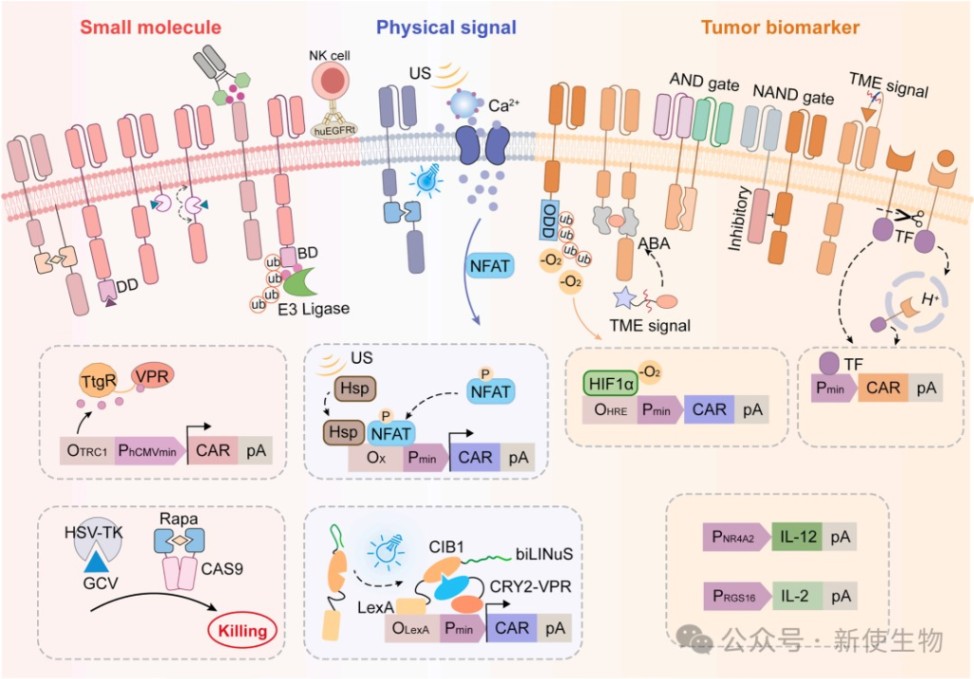
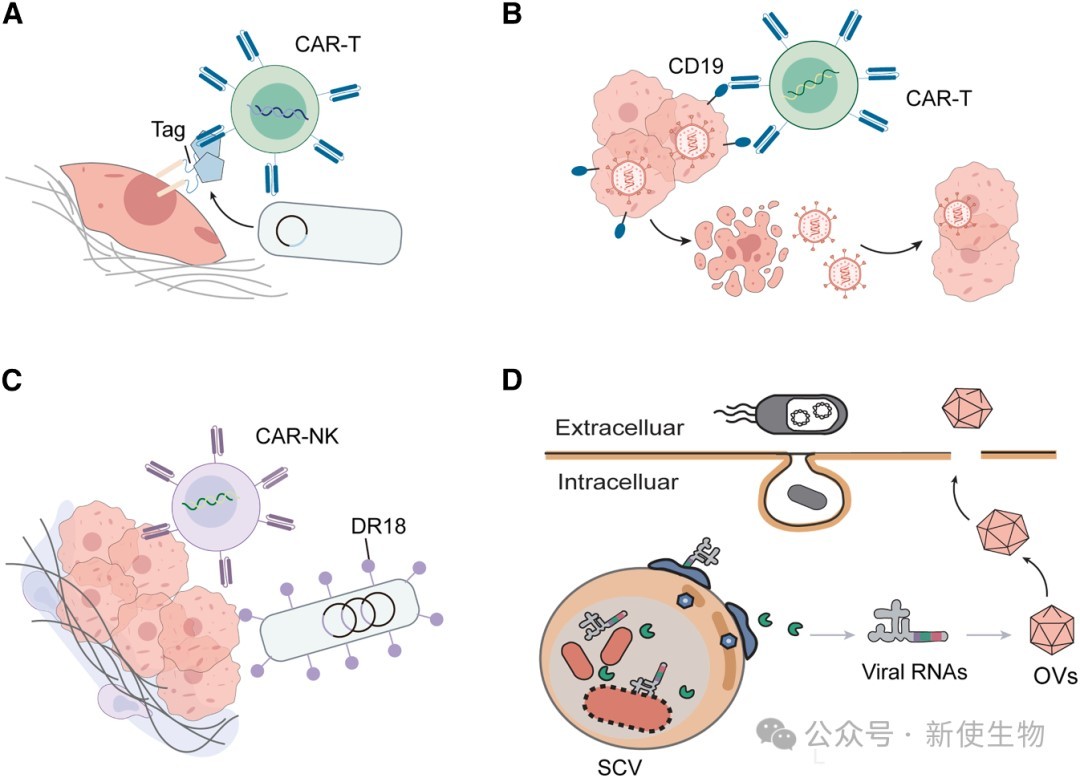
一、CAR-T细胞的合成基因开关
CAR-T细胞是目前临床最成熟、研究最广泛的哺乳动物底盘(Chassis)。
然而,过度激活引发的细胞因子释放综合征(CRS)、靶外毒性(On-target, off-tumor)以及T细胞耗竭严重制约了其应用。
合成生物学通过引入多维度的调节开关,显著提升了CAR-T的安全性与可控性。
1.1) 小分子诱导系统
该系统通过外部施用化学药物来实现对CAR-T活性的可逆与时间控制。
主要分为三类:
ON-开关:如依赖雷帕霉素或来那度胺诱导的CAR组装或表达。
OFF-开关:如利用小分子破坏蛋白相互作用或使用PROTAC技术降解CAR蛋白。
自杀开关:如引入iCasp9或HSV-TK,在出现严重毒性时诱导细胞凋亡。
目前,iCasp9等自杀开关在临床应用中最为深入。
1.2) 物理信号诱导系统
为了克服系统性给药带来的脱靶暴露和药代动力学差异,光和超声等物理信号被用于无创、时空受限的精准调控。
光遗传学模块能实现快速的光控激活,而基于Piezo1离子通道或热休克蛋白(Hsp)启动子的超声遗传学系统,为深层组织的精准激活提供了有效途径。
1.3) 肿瘤微环境(TME)诱导系统
TME响应型CAR-T无需外部干预,可根据肿瘤内的病理特征实现自主激活,设计策略包括:
1. 利用低氧诱导因子(HIF)结合氧依赖性降解(ODD)结构域实现低氧特异性表达。
2. 针对肿瘤酸性环境设计的pH敏感型CAR。
3. 开发能够感知TME中可溶性细胞因子(如VEGF、TGF-β)的合成受体(如SNIPR或MESA)。
4. 利用肿瘤相关蛋白酶裂解掩蔽肽来恢复CAR识别能力的“掩蔽CAR”(Masked CAR)。
1.4) 肿瘤抗原诱导系统
为了更精准地识别肿瘤细胞,研究者引入了布尔逻辑门控策略。
AND逻辑:如synNotch受体或LINK CAR要求目标细胞同时表达两种抗原才能激活T细胞,大幅降低了单抗原表达带来的脱靶毒性。
NOT逻辑:如iCAR通过融合PD-1或CTLA-4的抑制性信号域,在识别到正常组织抗原时阻止T细胞杀伤,从而保护健康组织。
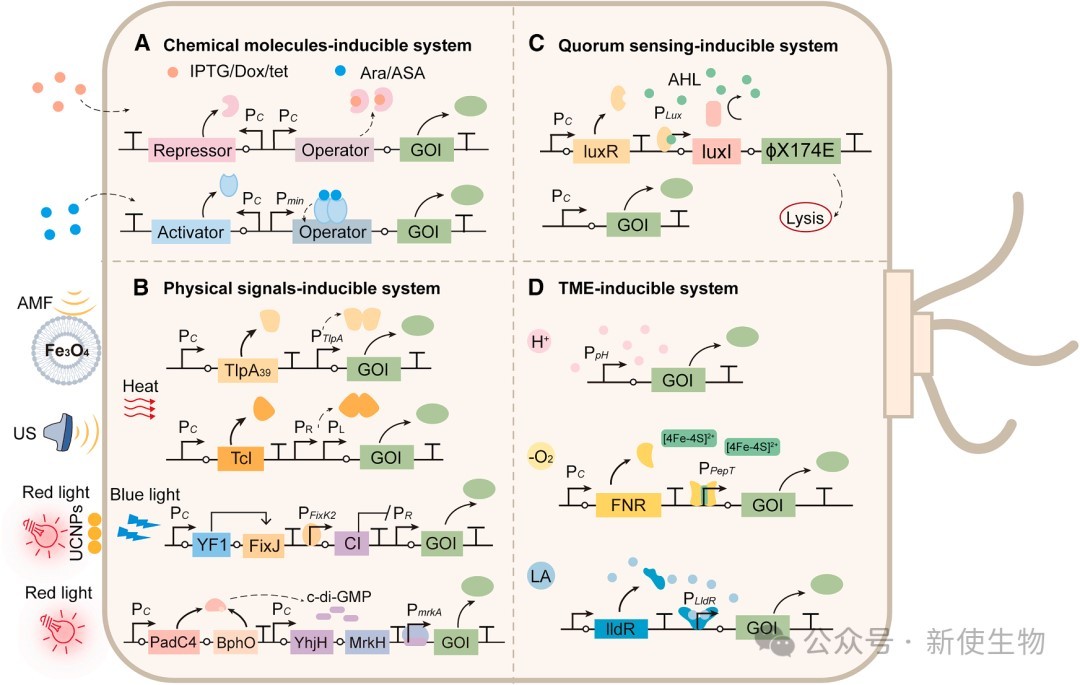
二、溶瘤细菌的程序化控制
部分厌氧或兼性厌氧细菌(如沙门氏菌、大肠杆菌Nissle 1917等)天然倾向于在肿瘤的缺氧和坏死区域定植,并在免疫抑制微环境中激活免疫反应。
通过合成基因回路对这些特性加以改造,可极大拓展其治疗潜力。
2.1) 小分子与物理信号调控
与CAR-T类似,细菌可通过小分子诱导系统(如IPTG、阿拉伯糖、多西环素或阿司匹林响应启动子)控制治疗蛋白的表达。
物理信号方面,结合上转换纳米颗粒或直接响应近红外光(NIR)的基因回路(如NETMAP系统),以及基于温度敏感型阻遏蛋白的超声响应系统,实现了对细菌定植、裂解和载荷释放的无创控制。
2.2) 群体感应(QS)系统
细菌可通过群体感应信号(如AHL)进行自发通信,研究人员利用该机制设计了同步裂解回路(SLCs)。
当肿瘤内的细菌密度达到临界阈值时,自动触发细菌裂解并向TME释放预先表达的治疗载荷(如趋化因子、毒素或免疫检查点纳米抗体),这为局部抗癌药物递送提供了一种高度自主的范式。
2.3) TME响应与逻辑门控
细菌可通过感知低氧(利用FNR调节器)、低pH值或高乳酸浓度来实现肿瘤选择性激活。
为进一步提升特异性,研究构建了多输入逻辑门,确保仅在特定TME特征组合出现时才释放杀伤性蛋白。
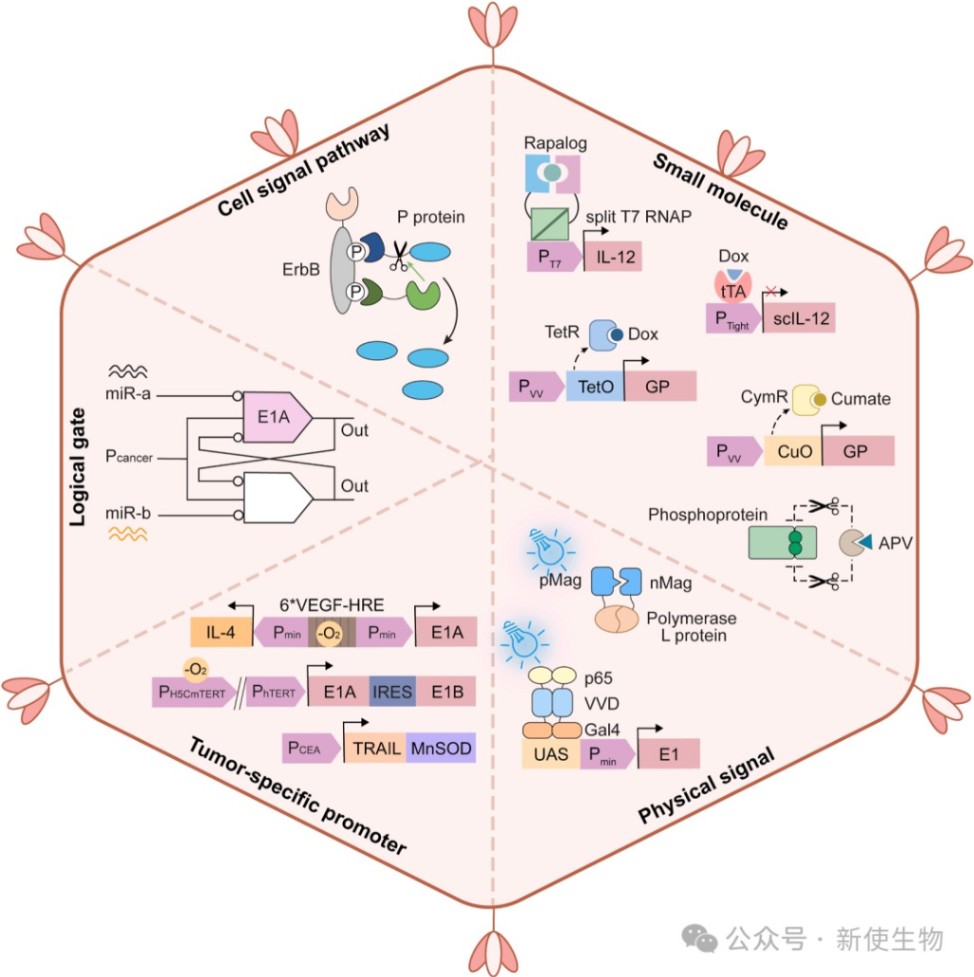
三、溶瘤病毒的工程化改造
溶瘤病毒(OVs)能够选择性地感染肿瘤细胞并使其裂解,同时引发全身性抗肿瘤免疫。
为了克服天然病毒靶向性不足及易引发抗病毒免疫的缺陷,合成生物学对其进行了深度重编程。
3.1) 化学与物理信号诱导
通过将雷帕霉素、多西环素或蛋白酶抑制剂的响应元件嵌入病毒基因组,可精确调控治疗基因的表达或病毒的复制。
光遗传学系统(如蓝光调控的聚合酶L蛋白或E1基因)也被成功用于精准激活病毒的溶瘤活性。
3.2) 肿瘤特异性信号响应
这是目前在临床上进展最快的策略。通过使用肿瘤特异性启动子(如hTERT、AFP或HIF响应启动子)替代天然启动子,可限制病毒仅在癌细胞中复制。
此外,将健康细胞中高表达的microRNA(如miR-122)的靶序列插入病毒基因组,可有效阻止病毒在正常组织中的复制。
此外,利用癌细胞中缺陷的信号通路(如p53、Rb或高活性的Ras通路)也是增强肿瘤特异性的常用手段。
3.3) 病毒表面蛋白重定向
通过重编程病毒表面蛋白以改变其趋向性(Tropism),可最小化脱靶感染。
具体方法包括使用双特异性单链抗体、工程化锚蛋白重复蛋白(DARPins)或在糖蛋白上展示特定scFv(如靶向HER2或CD20),引导病毒特异性地结合肿瘤表面受体。
四、临床转化的挑战与底盘特异性限制
4.1) 各底盘的转化壁垒
不同底盘的生物学特性决定了其临床转化的难度。
CAR-T细胞:受限于高昂的制造成本和基因组包装容量,倾向于采用紧凑的药物门控开关。
细菌:能够承载复杂的逻辑门,但面临高突变率、进化不稳定性及质粒水平转移等生物安全隐患。
溶瘤病毒:受制于极小的有效载荷容量和维持复制适应性的需求,通常只能采用极简的调控元件(如特异性启动子或miRNA靶点)。
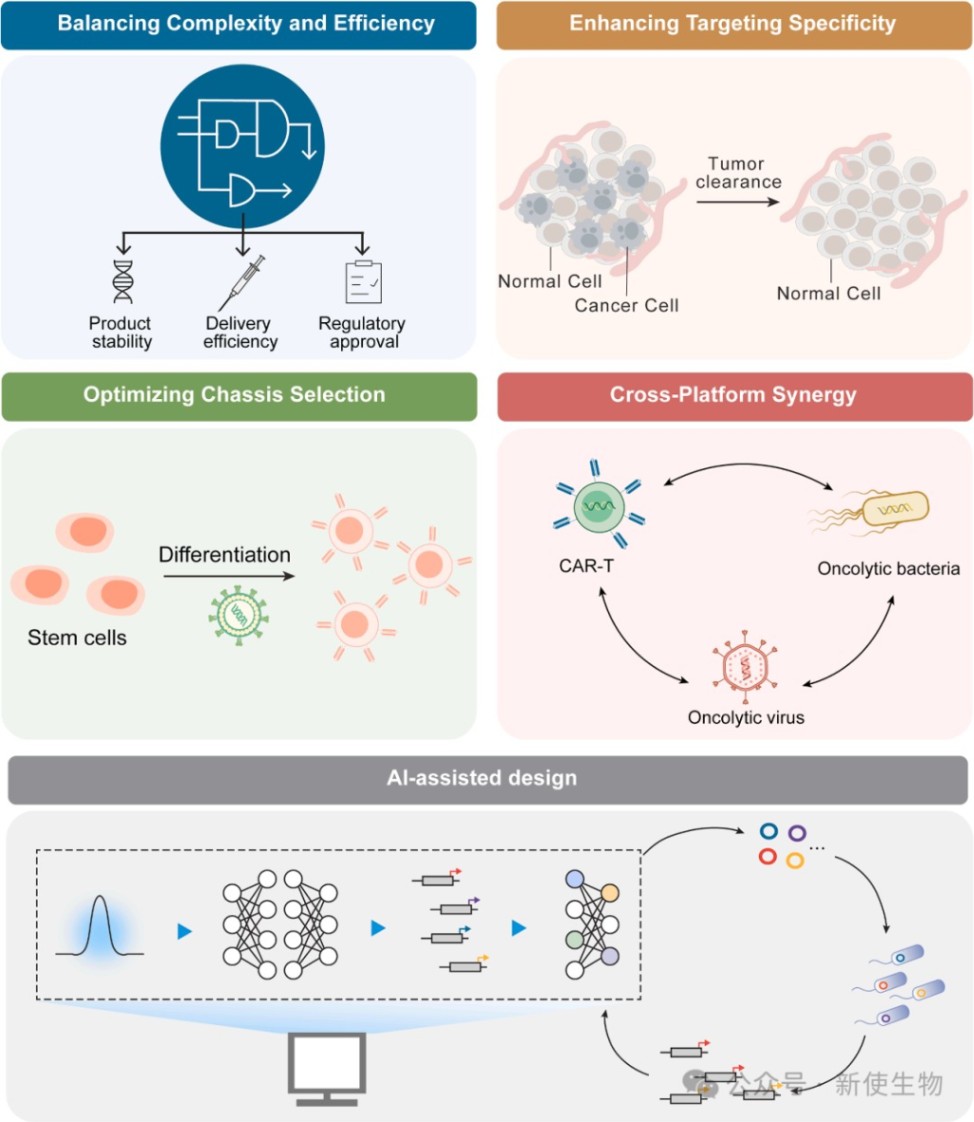
4.2) 基因稳定性与生物安全
活体药物在体内选择压力下易发生突变、缺失或表观遗传沉默。
对于溶瘤病毒,重组风险可能导致其恢复毒力,而对于细菌,则需引入营养缺陷型或多重生物安全壳来防止环境泄漏。
未来设计必须将底盘感知(Chassis-aware)作为核心原则,平衡回路复杂性与进化稳健性。
总结与展望
可编程的活体疗法已超越传统的固定生化药物,成为具有“感知-计算-执行”能力的下一代癌症免疫治疗核心。然而,不同shw底盘在基因组容量、代谢负担及进化稳定性上具有显著的特异性限制。
未来的临床转化不仅需要简化基因线路以提高遗传稳定性和制造可行性,还需要拥抱生成式人工智能(AI)辅助的序列设计。此外,CAR-T、细菌与病毒的跨平台协同将开启多维度协同抗癌的新纪元。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看