

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
传统的蛋白质编码基因注释依赖于序列保守性、AUG起始密码子和长度大于100个密码子等标准。然而,核糖体印迹分析(Ribo-seq)技术的进步揭示了基因组中存在大量非经典翻译事件,它们主要发生在信使RNA的非翻译区(UTR)或非编码RNA上。
这些翻译事件产生了成千上万种通常被称为微蛋白的新型多肽。尽管微蛋白在转录和翻译水平上广泛存在,但它们在蛋白质水平的丰度却极低,其生物学功能也大多未知。
有趣的是,源自微蛋白的多肽被广泛地呈递在人类白细胞抗原(HLA)分子上,暗示它们可能在免疫调节中发挥作用。这种翻译水平与蛋白质丰度之间的巨大差异表明,大多数微蛋白可能在翻译后被迅速降解,但其背后的分子机制和生物学逻辑尚不完全清楚。
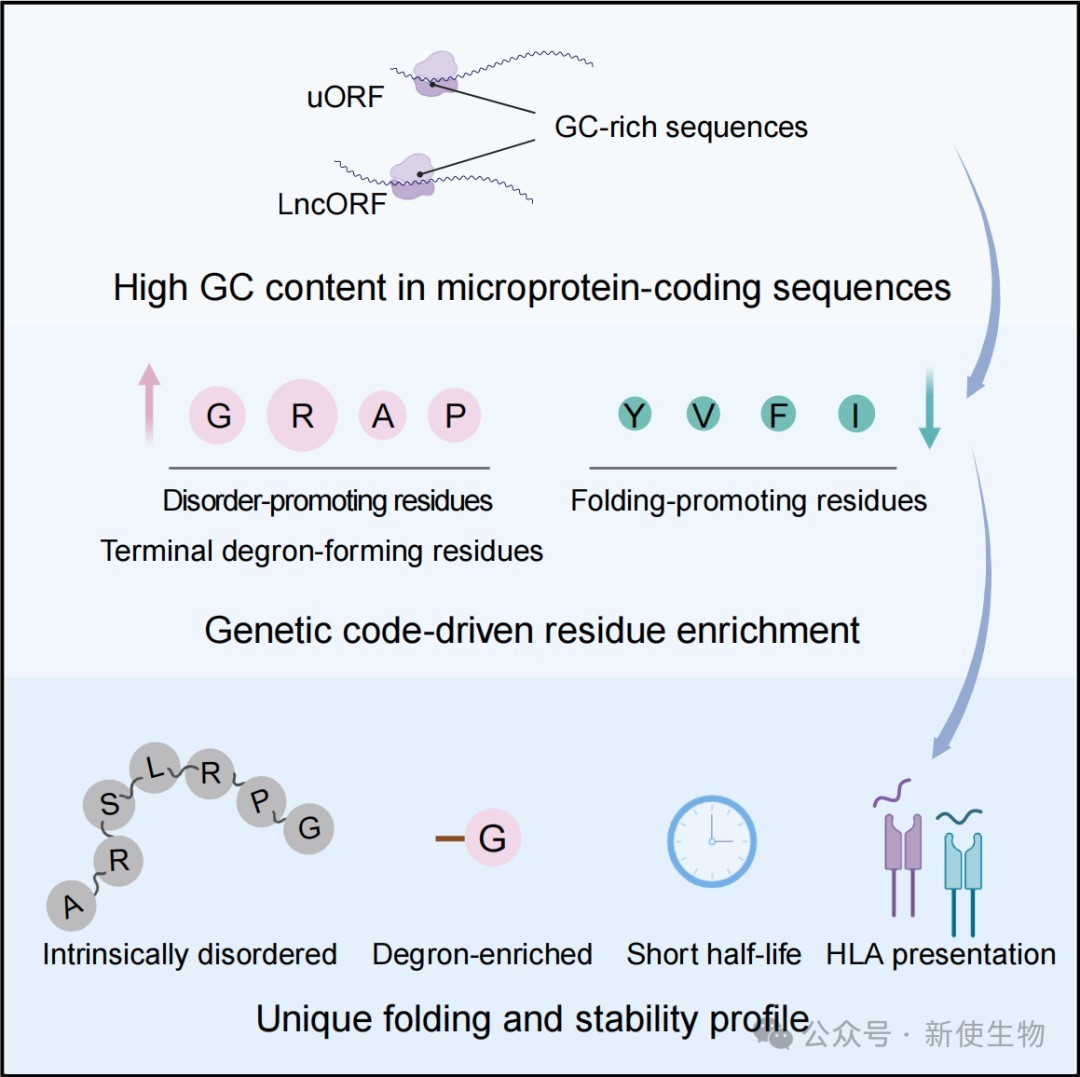
2026年5月20日,浙江大学张汕和王勇团队合作在Molecular Cell上发表了题为“Intrinsic bias of the genetic code shapes the folding and stability landscapes of microproteins” 的研究论文。该研究揭示了人类微蛋白折叠和稳定性的基本规律,即编码序列的高GC含量与遗传密码的内在偏好性共同塑造了微蛋白的氨基酸组成。

文章索引
【标题】Intrinsic bias of the genetic code shapes the folding and stability landscapes of microproteins
【发表期刊】Molecular Cell
【发表日期】2026年5月20日
【作者及团队】浙江大学张汕和王勇团队
【IF】16.6
研究结果
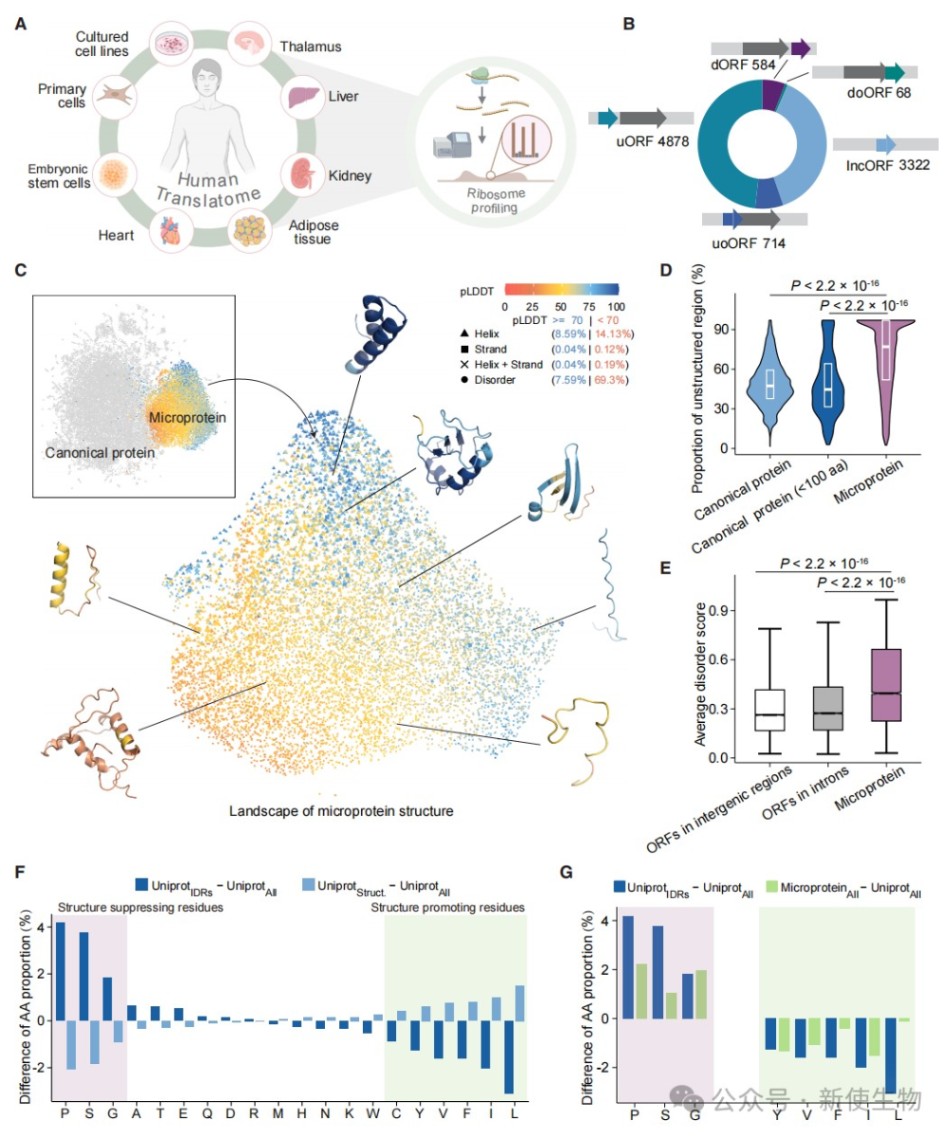
一、微蛋白普遍呈现内在无序且折叠倾向极低
为鉴定人类微蛋白,研究人员整合了来自20多种细胞和组织的高质量Ribo-seq核糖体印迹分析数据,共识别出9,566个被翻译的非经典开放阅读框(nORF)。
研究通过ESMFold和AlphaFold2结构预测发现,绝大多数微蛋白缺乏稳定的二级结构,表现出高度的内在无序性。
这种结构特征主要源于其氨基酸组成的偏好,即富含脯氨酸、丝氨酸、甘氨酸和丙氨酸等结构抑制性残基。
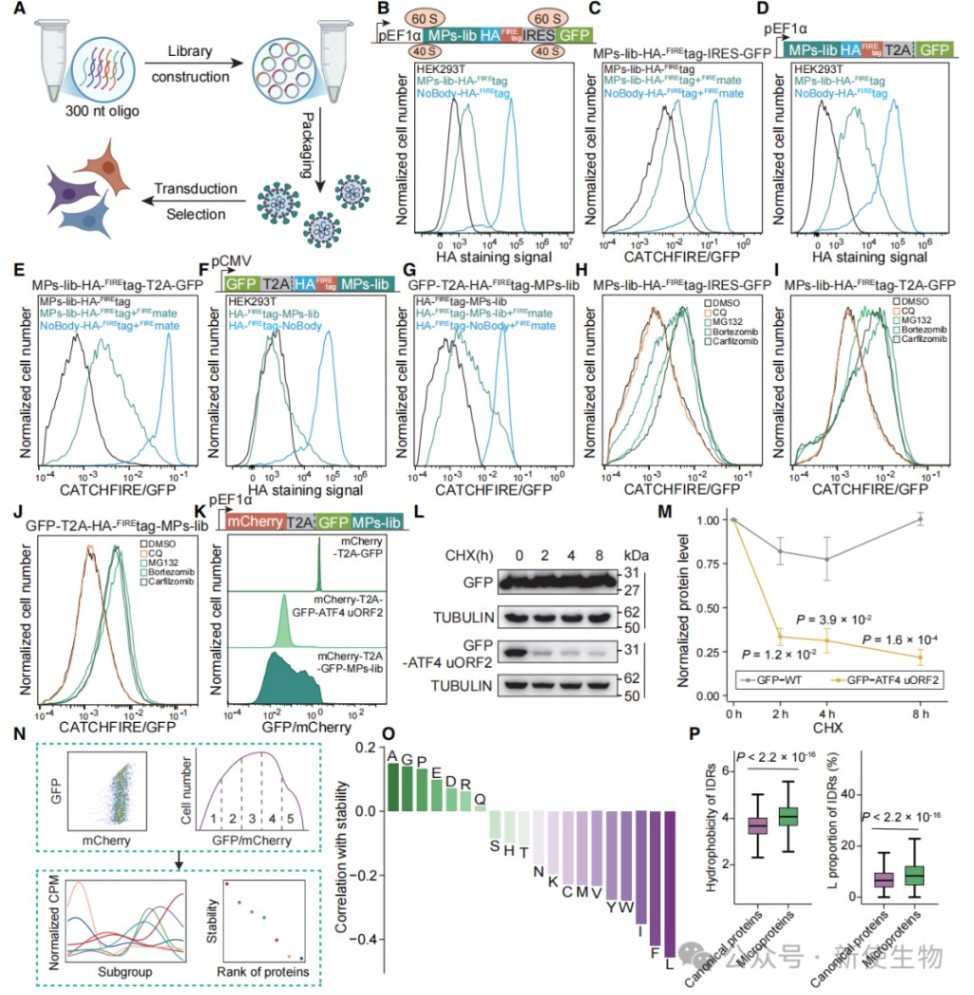
二、微蛋白通过泛素-蛋白酶体途径被快速降解
利用超长寡核苷酸库构建的微蛋白表达系统显示,微蛋白在细胞内的稳态水平极低。
通过化学抑制剂筛选和动力学分析,证实微蛋白主要通过泛素-蛋白酶体系统而非溶酶体途径进行快速降解。
三、末端降解信号是微蛋白被识别的关键
全基因组CRISPR筛选鉴定出CUL2等E3泛素连接酶复合物负责识别微蛋白的末端降解信号。
研究发现,C端甘氨酸和特定位置的精氨酸等残基构成了强效的降解基序,突变这些基序可显著提高微蛋白的稳定性。
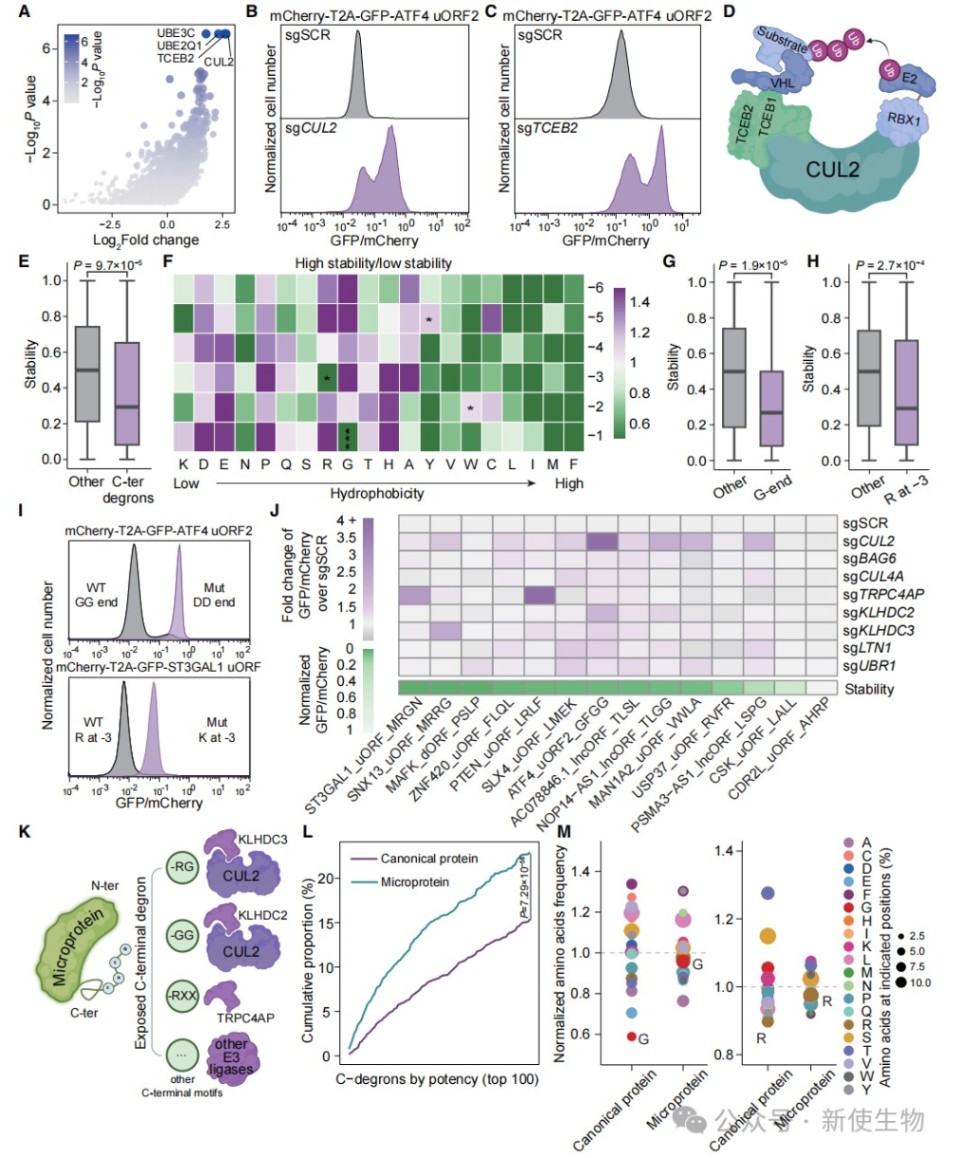
四、疏水性与末端降解信号共同驱动微蛋白的不稳定性
通过构建双荧光报告系统,量化了疏水性残基与末端降解信号对稳定性的影响。
实验表明,约80%的微蛋白不稳定性可归因于这两类机制的协同作用,且BAG6途径仅在极少数高疏水性微蛋白中发挥作用。
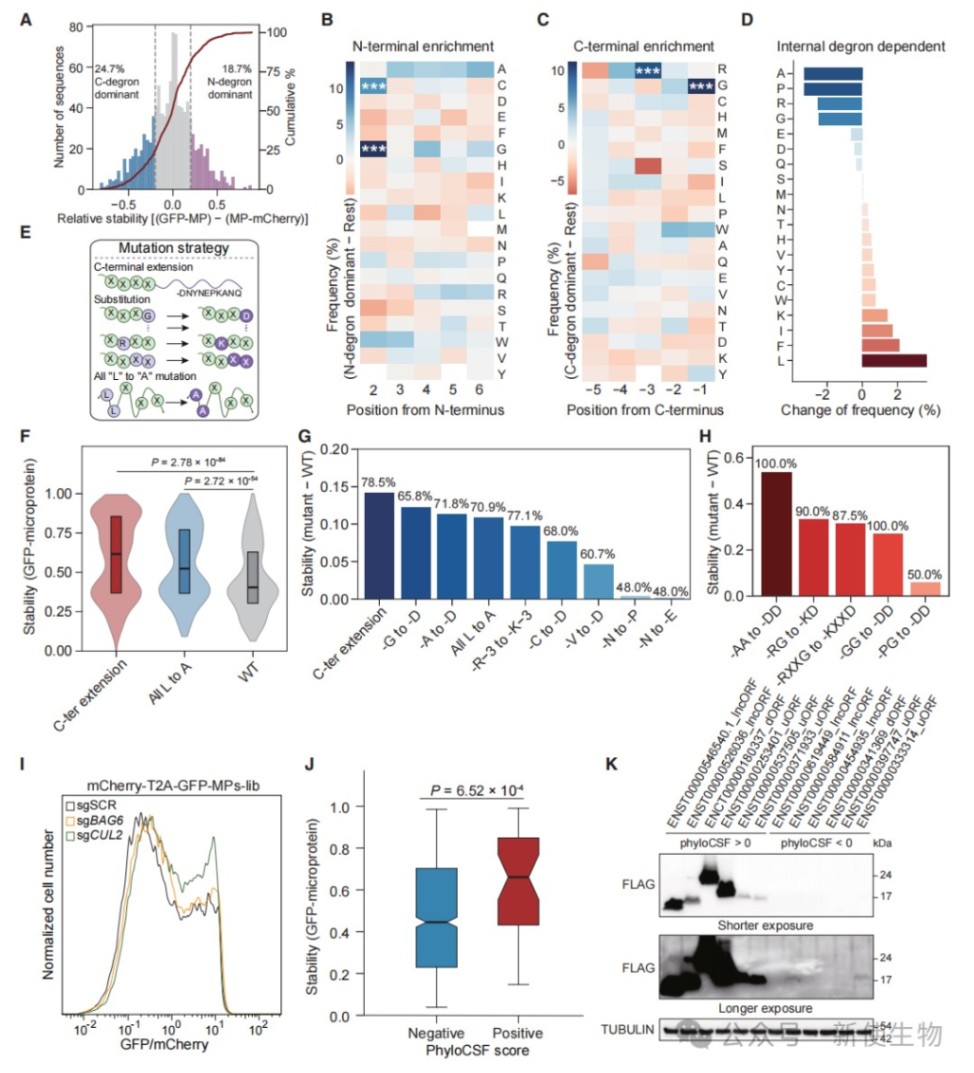
五、遗传密码偏好与高GC含量塑造微蛋白组成
微蛋白编码序列的高GC含量导致其富含由GC丰富密码子编码的残基,即甘氨酸、精氨酸、丙氨酸和脯氨酸。
这种遗传密码的内在偏好性解释了微蛋白为何在进化上倾向于形成无序且易降解的序列。
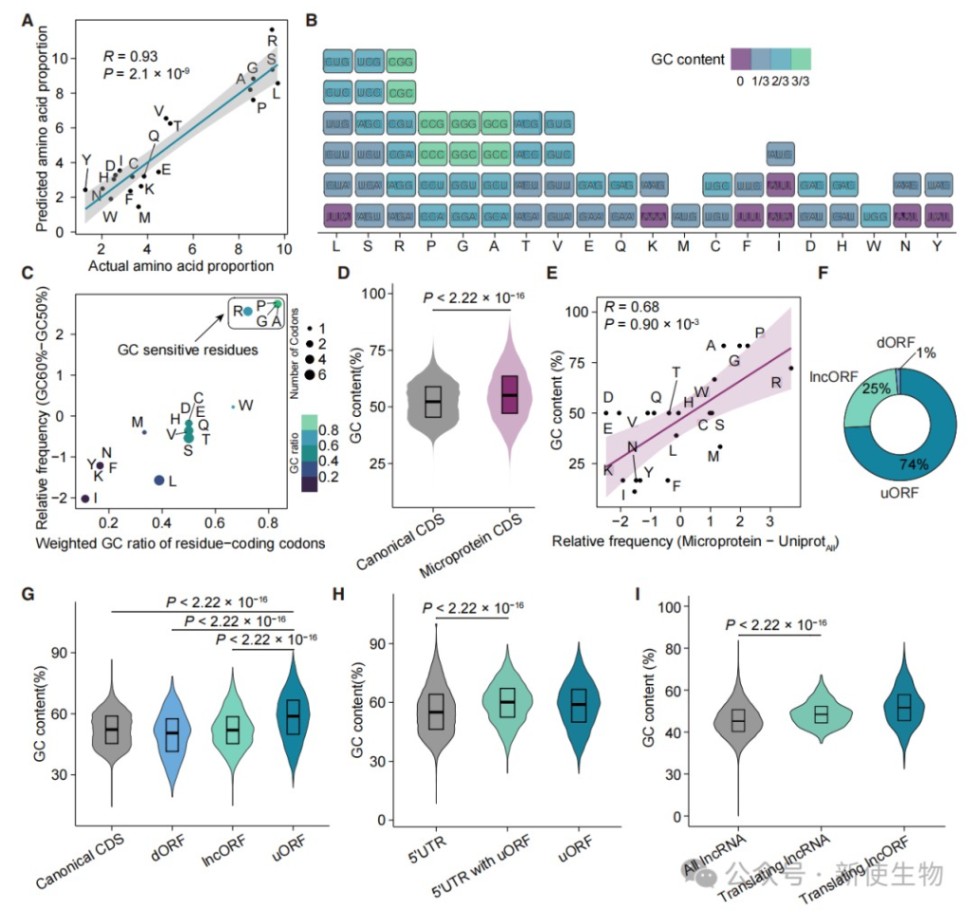
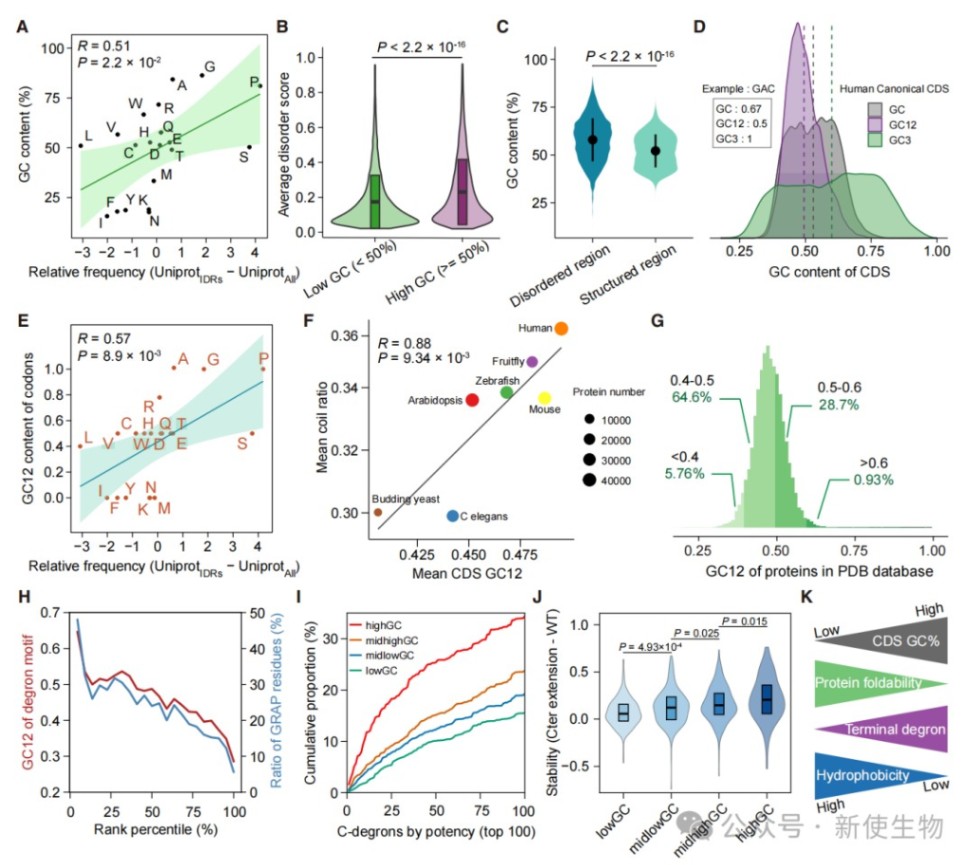
六、GC含量是决定蛋白质折叠与质量控制的根本因素
分析表明,编码序列的GC12指数与蛋白质的无序程度呈强正相关,且该规律在不同物种间高度保守。
高GC含量不仅导致蛋白质折叠困难,还通过增加末端降解信号的频率,直接决定了蛋白质的质量控制命运。
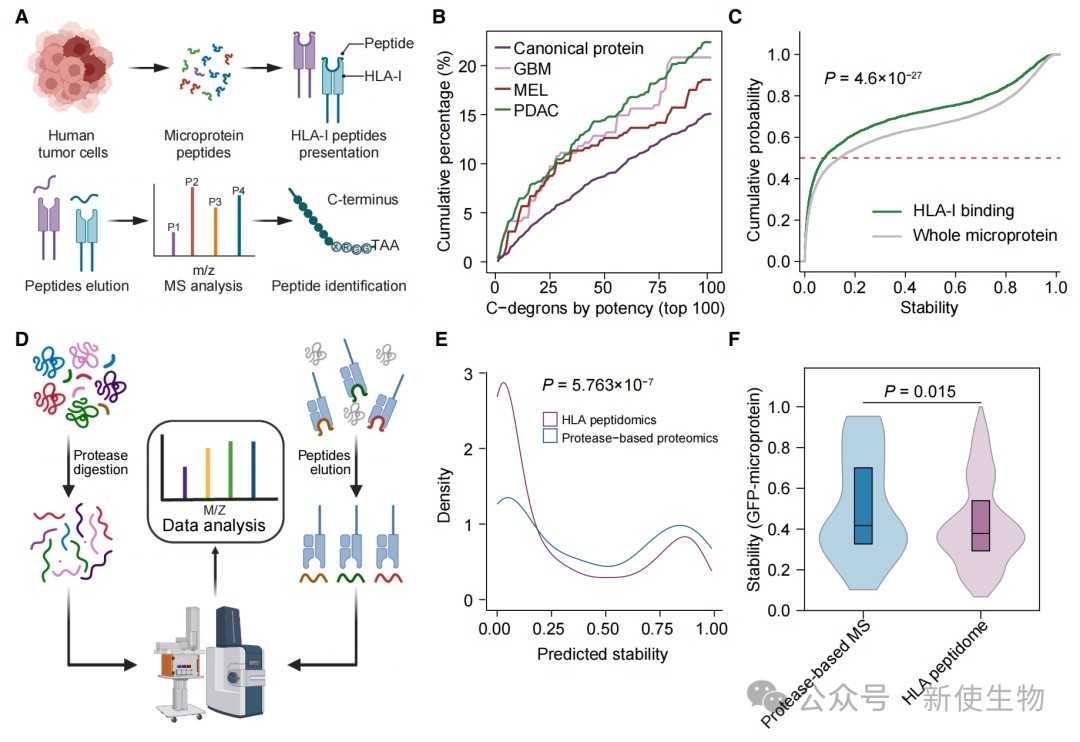
七、不稳定微蛋白是免疫肽组的重要来源
研究发现,被HLA-I分子呈递的微蛋白肽段显著富含末端降解信号。
通过神经网络模型预测证实,那些被快速降解的微蛋白片段更易进入抗原呈递途径,从而成为肿瘤免疫监视的重要靶点。
总结
本研究建立了一个简洁的定量模型,揭示了遗传密码的内在偏好性如何通过GC含量影响微蛋白的折叠与稳定性。该机制不仅解释了微蛋白为何普遍不稳定,还阐明了细胞如何利用这一特性将非经典翻译产物转化为免疫呈递的抗原库。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看