

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
人类线粒体基因组编码13种氧化磷酸化(OXPHOS)系统的核心亚基,对细胞能量代谢至关重要。这些蛋白的翻译在线粒体内膜(IMM)上进行,并与新生肽链通过OXA1L插入酶共翻译插入膜内的过程相偶联。
新合成的肽链随后与核编码的亚基组装,共同构建功能性的OXPHOS复合体。然而,新生肽链膜插入的具体机制,以及该过程与翻译机器之间的功能联系尚不明确。
尽管近期的冷冻电镜研究揭示了线粒体核糖体与OXA1L复合物的结构,但仍缺乏实时动态信息来解释翻译合成与膜插入的协同作用。此外,线粒体翻译过程中核糖体暂停的生物学意义,以及多跨膜结构域蛋白的插入机制等关键问题仍有待解答。
2026年5月7日,德国哥廷根大学Peter Rehling团队与马普多学科科学研究所Niels Fischer团队合作,在Nature Structural & Molecular Biology上发表了题为“Membrane insertion of mitochondrial-encoded proteins regulates ribosome decoding speed”的研究论文。该研究通过结合核糖体印迹分析(Ribo-seq)与冷冻电镜技术,发现线粒体新合成多肽的膜拓扑结构会显著影响核糖体的翻译速度。

文章索引
【标题】Membrane insertion of mitochondrial-encoded proteins regulates ribosome decoding speed
【发表期刊】Nature Structural & Molecular Biology
【发表日期】2026年5月7日
【作者及团队】德国哥廷根大学Peter Rehling和马克斯·普朗克多学科科学研究所Niels Fischer 团队
【IF】10.1
研究结果
一、建立并验证线粒体核糖体足迹分析技术
研究人员优化了适用于线粒体翻译的Ribo-seq核糖体印迹分析流程,通过氯霉素快速阻断翻译并利用微球菌核酸酶消化,成功分离并测序了30-40nt的线粒体核糖体保护片段。
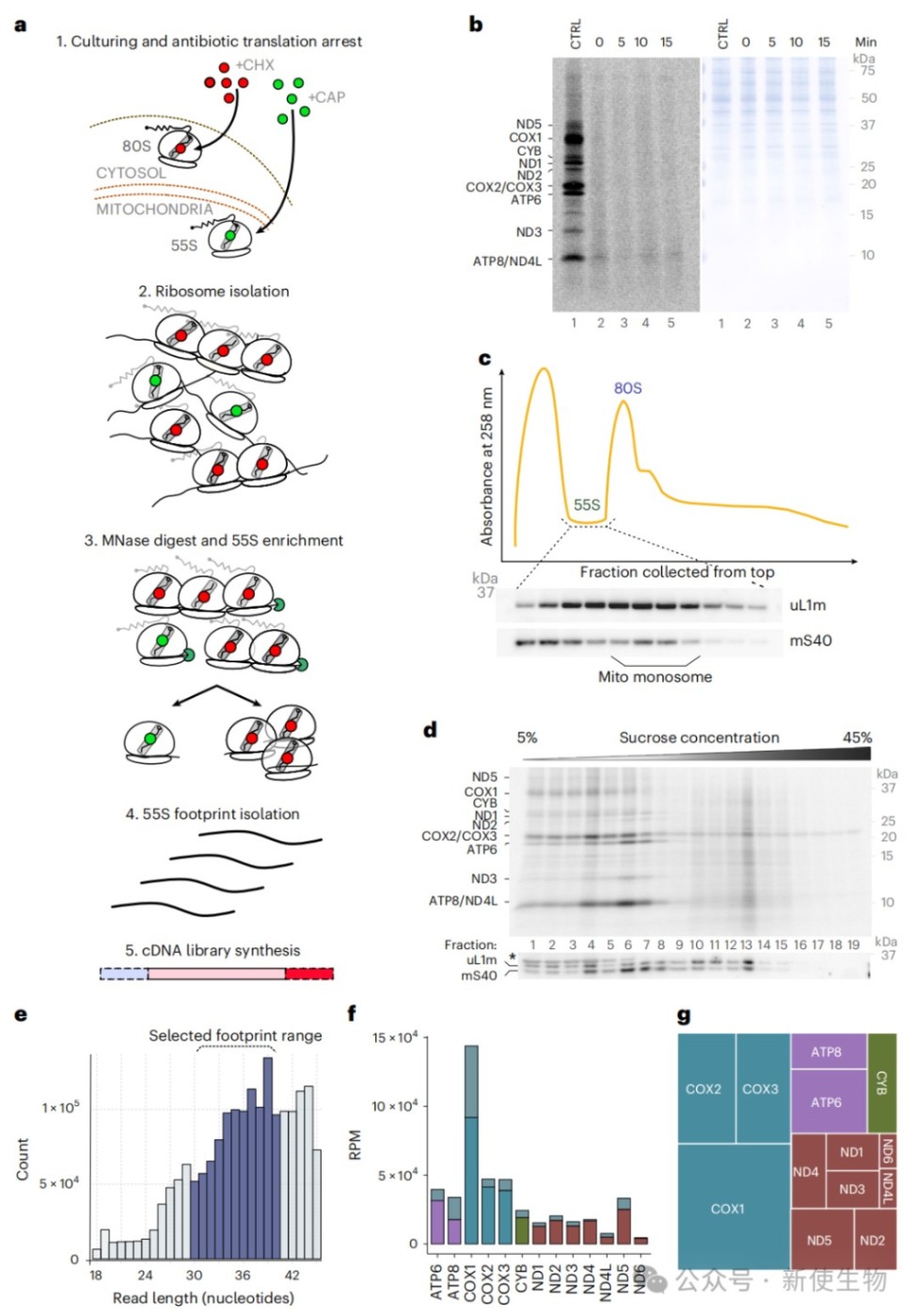
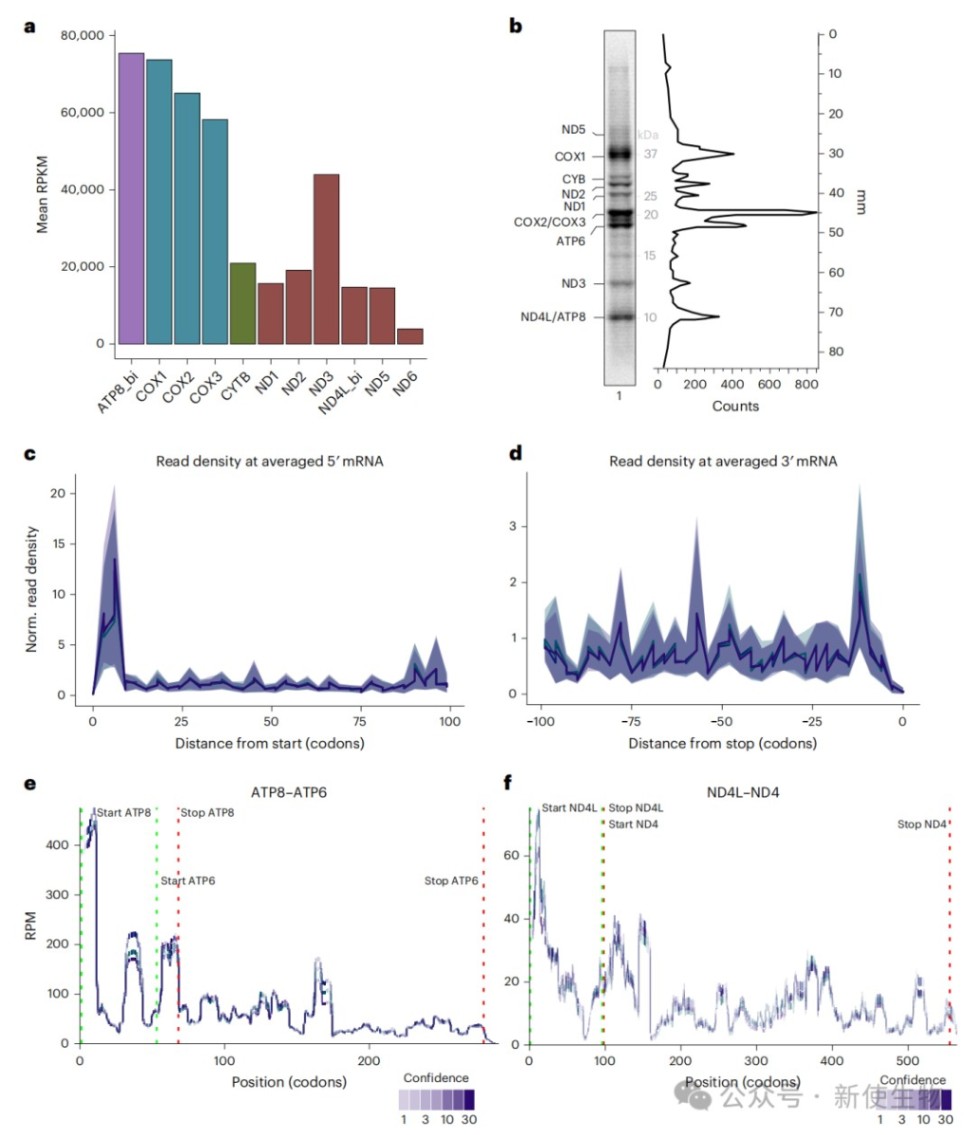
数据分析表明,该方法能够精确绘制所有13种线粒体编码mRNA上的核糖体分布图谱,其读段丰度与放射性同位素标记的翻译产物水平高度一致。
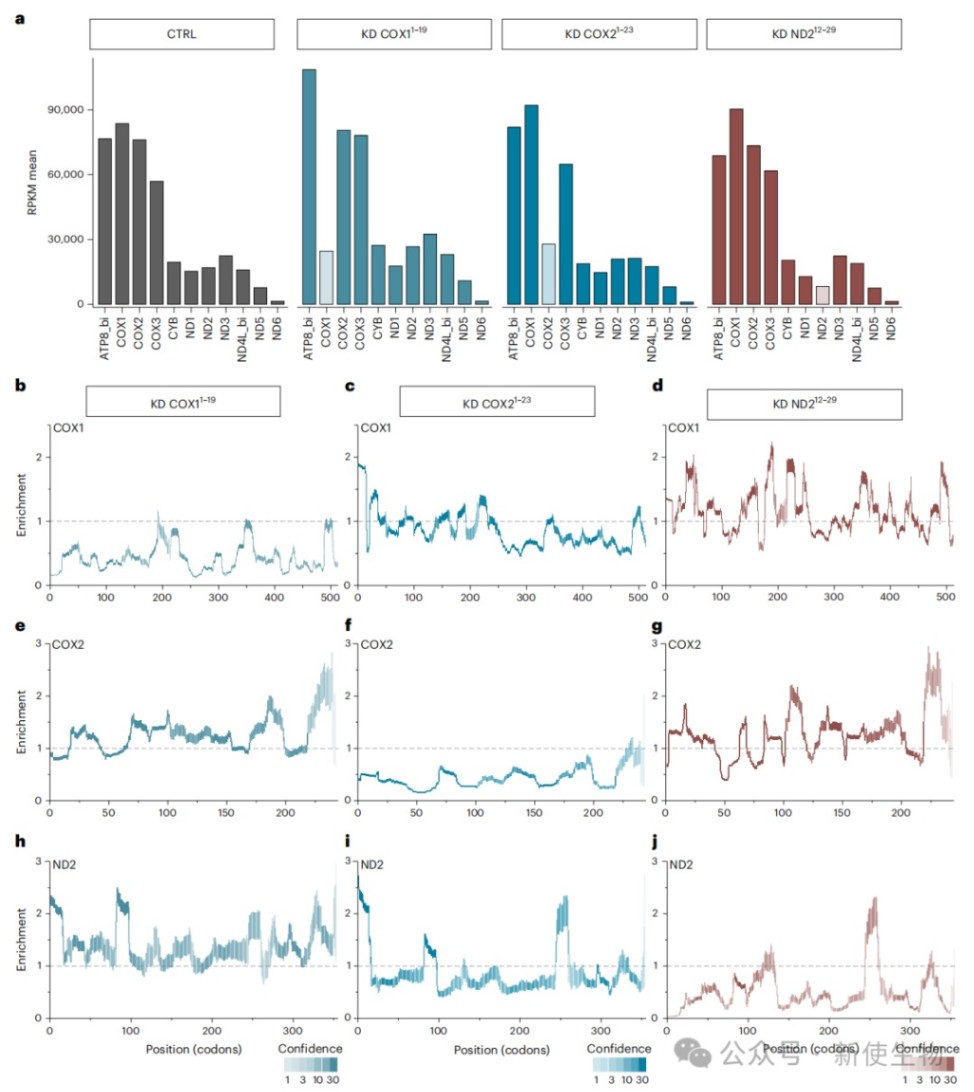
二、细胞器内基因沉默特异性降低靶mRNA的核糖体密度
利用导入线粒体的多聚吗啉代寡核苷酸特异性沉默COX1、COX2或ND2的翻译,核糖体足迹分析显示靶mRNA上的核糖体占位显著降低。
然而,阻断COX2翻译还会引起其他非靶标线粒体转录本翻译动力学的复杂变化,验证了该技术在监测线粒体基因表达调控中的高灵敏度。
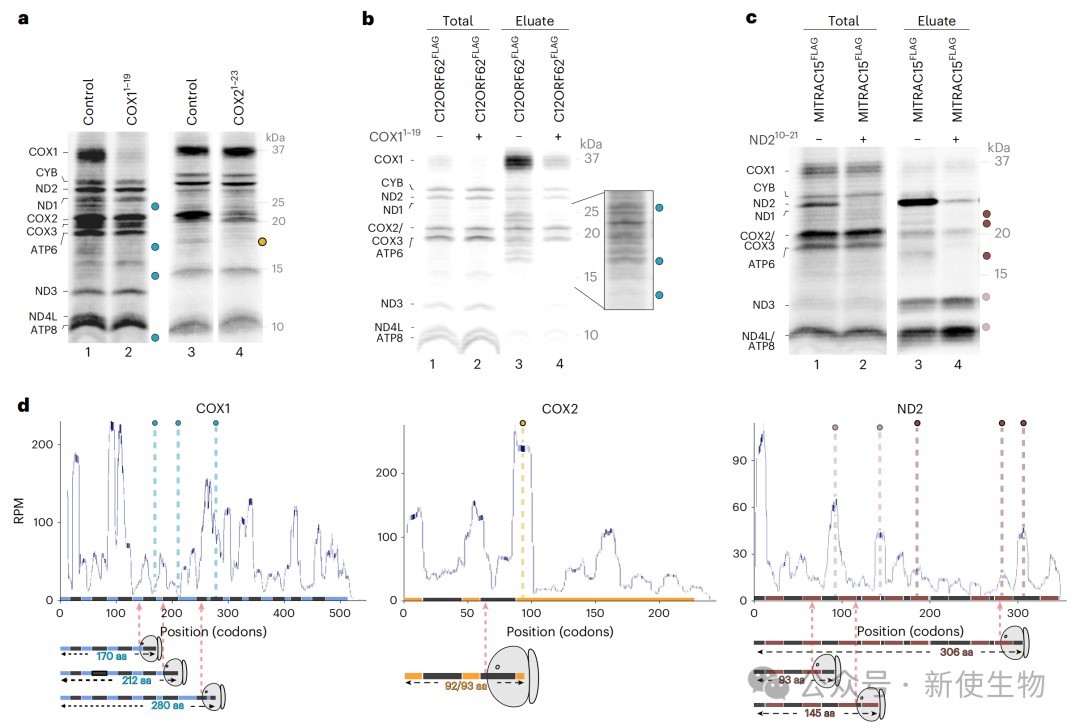
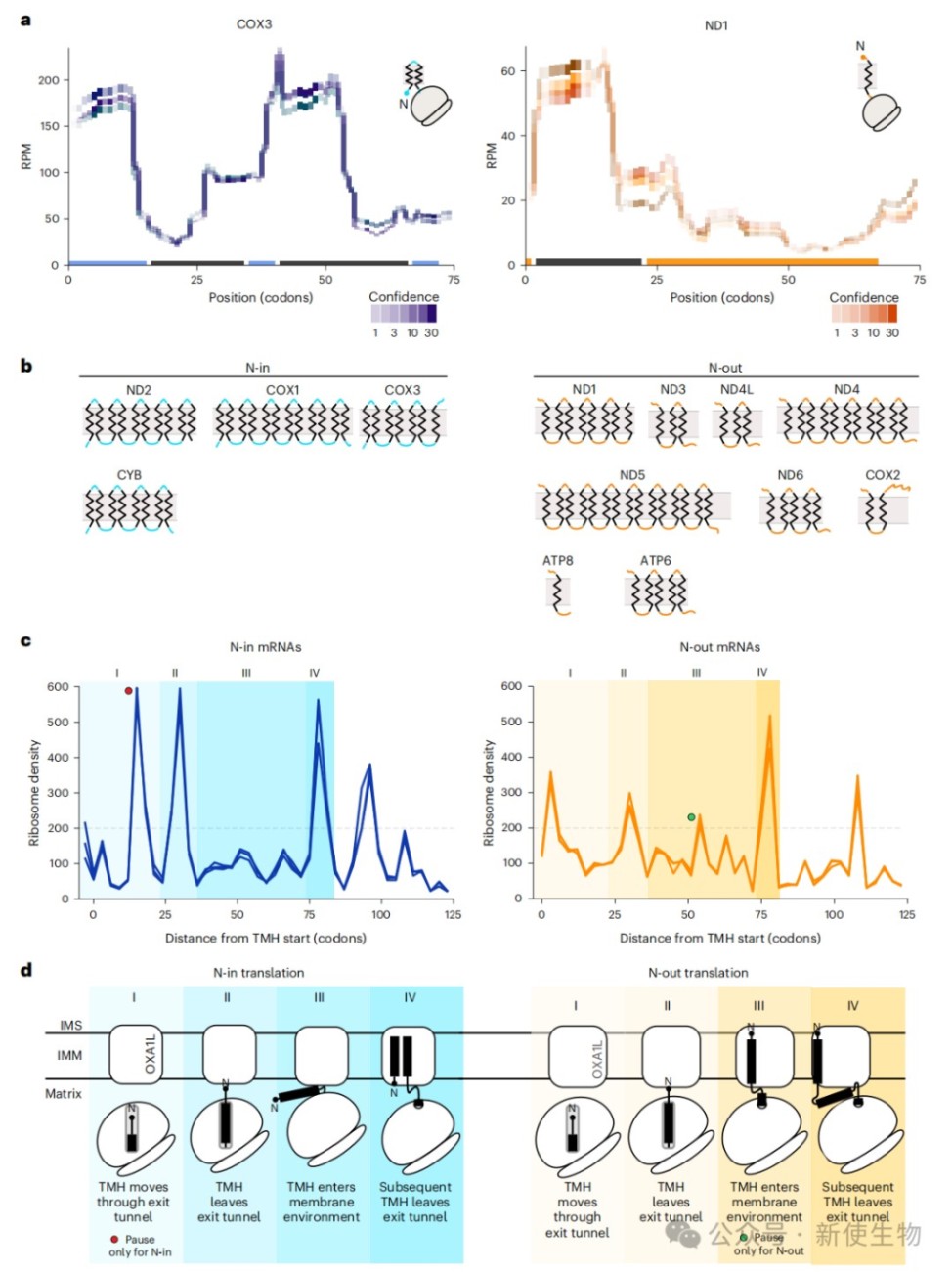
三、核糖体翻译暂停与跨膜螺旋的膜整合密切相关
通过对比细胞器内翻译产生的新生肽链与Ribo-seq核糖体印迹分析结果,研究人员发现COX1、COX2和ND2在翻译过程中存在特定的暂停位点。
这些暂停事件高度富集于跨膜螺旋刚离开核糖体出口通道或前庭的时刻,表明翻译速度会根据膜整合的需要进行动态调整。
四、新生肽链的膜拓扑结构决定核糖体解码速度
根据N端定位将线粒体膜蛋白分为N端在基质(N-in)和N端在膜间隙(N-out)两类,元基因分析显示两者的核糖体暂停模式存在显著差异。
N-in蛋白在翻译早期表现出特异性的暂停,表明在核糖体出口前庭形成发卡结构以维持正确的膜拓扑会显著降低翻译延伸速度。
五、冷冻电镜揭示COX1共翻译折叠与核糖体前庭的动态偶联
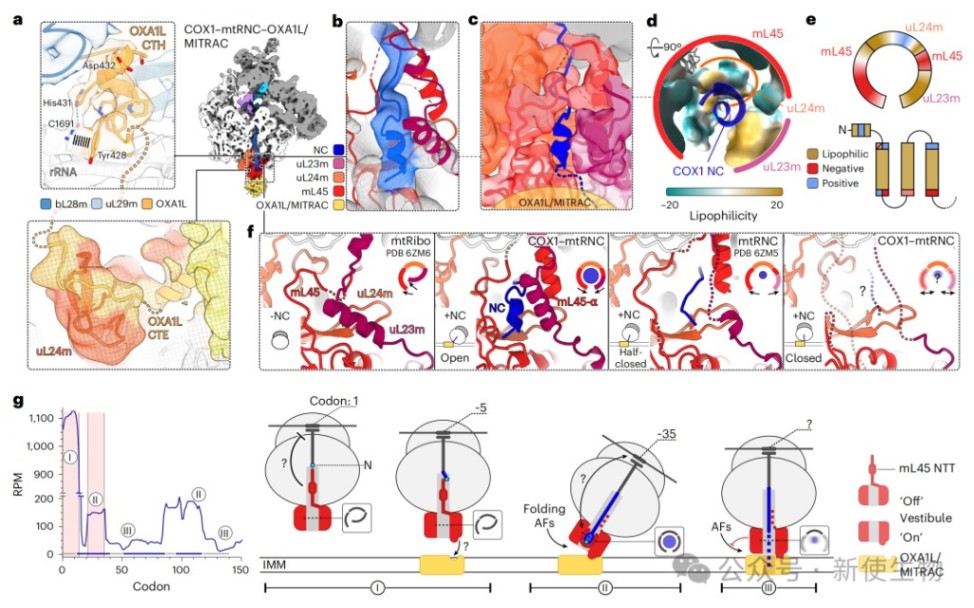
研究人员解析了处于不同翻译阶段的COX1–mtRNC–OXA1L/MITRAC复合物高分辨率结构,首次观察到COX1新生链在核糖体前庭内折叠成双亲性α螺旋。
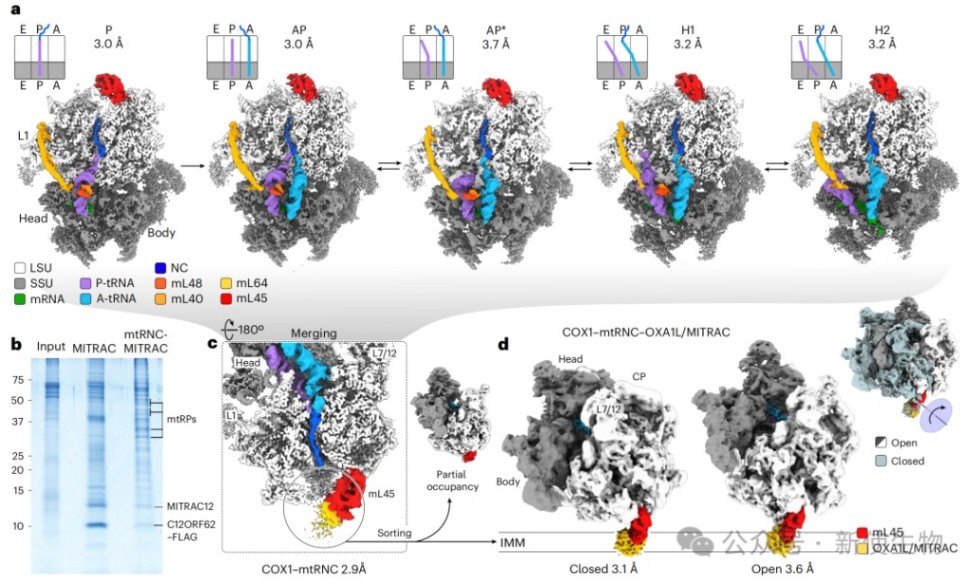
这种共翻译折叠诱导了前庭的局部构象改变,并与复合物整体的开放和关闭状态旋转相关联,从而在空间上调节辅助因子的结合与新生链的膜插入。
总结
本研究结合核糖体分析和冷冻电镜技术,系统揭示了人类线粒体中蛋白质的共翻译膜插入过程如何动态调控核糖体的解码速度。研究发现新生肽链的膜拓扑结构和两亲性螺旋的折叠会诱导翻译暂停,为理解线粒体基因表达与OXPHOS系统生物合成的协同调控提供了关键的分子机制。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看