

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
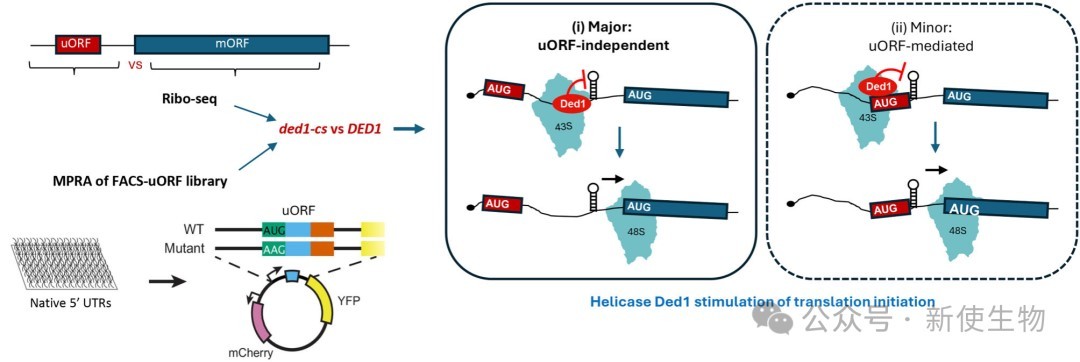
真核生物的翻译起始主要通过扫描机制进行,其中43S前起始复合物结合到mRNA的5'端并沿5' UTR寻找起始密码子。在这一过程中,5' UTR内的二级结构会显著阻碍复合物的结合与扫描。
为了克服这些障碍,细胞需要借助DEAD-box家族的RNA解旋酶(如eIF4A和Ded1/DDX3)来解开这些复杂的二级结构。过去的研究表明,在多聚核糖体组装受阻的情况下,Ded1对于翻译具有高度结构化5' UTR的mRNA依然至关重要。
此外,位于微弱起始密码子下游的二级结构有时会通过增加核糖体的停留时间来促进该位点的起始,这种机制被称为结构辅助的RNA翻译。曾有模型提出,Ded1主要通过解开uORF下游的二级结构来抑制这些上游开放阅读框的翻译,从而间接促进下游主开放阅读框的翻译。
近日,美国国立卫生研究院Alan G. Hinnebusch团队在Nucleic Acids Research上发表了题为“Suppression of upstream ORF translation is not a widespread mechanism of translational stimulation by yeast helicase Ded1” 的研究论文。该研究发现酵母解旋酶Ded1对翻译起始的刺激作用,主要通过解旋5' UTR二级结构本身来实现,而不是广泛地通过抑制上游开放阅读框(uORF)的翻译。

 |
文章索引
【标题】Suppression of upstream ORF translation is not a widespread mechanism of translational stimulation by yeast helicase Ded1
【发表期刊】Nucleic Acids Research
【发表日期】2026年4月11日
【作者及团队】美国国立卫生研究院Alan G. Hinnebusch团队
【IF】13.1
研究结果
一、ded1突变体中下游mORF翻译降低并非由uORF翻译增加引起
研究人员在不使用环己酰亚胺处理细胞的改良条件下进行了Ribo-seq核糖体印迹分析,以避免人工造成的5' UTR核糖体占位。
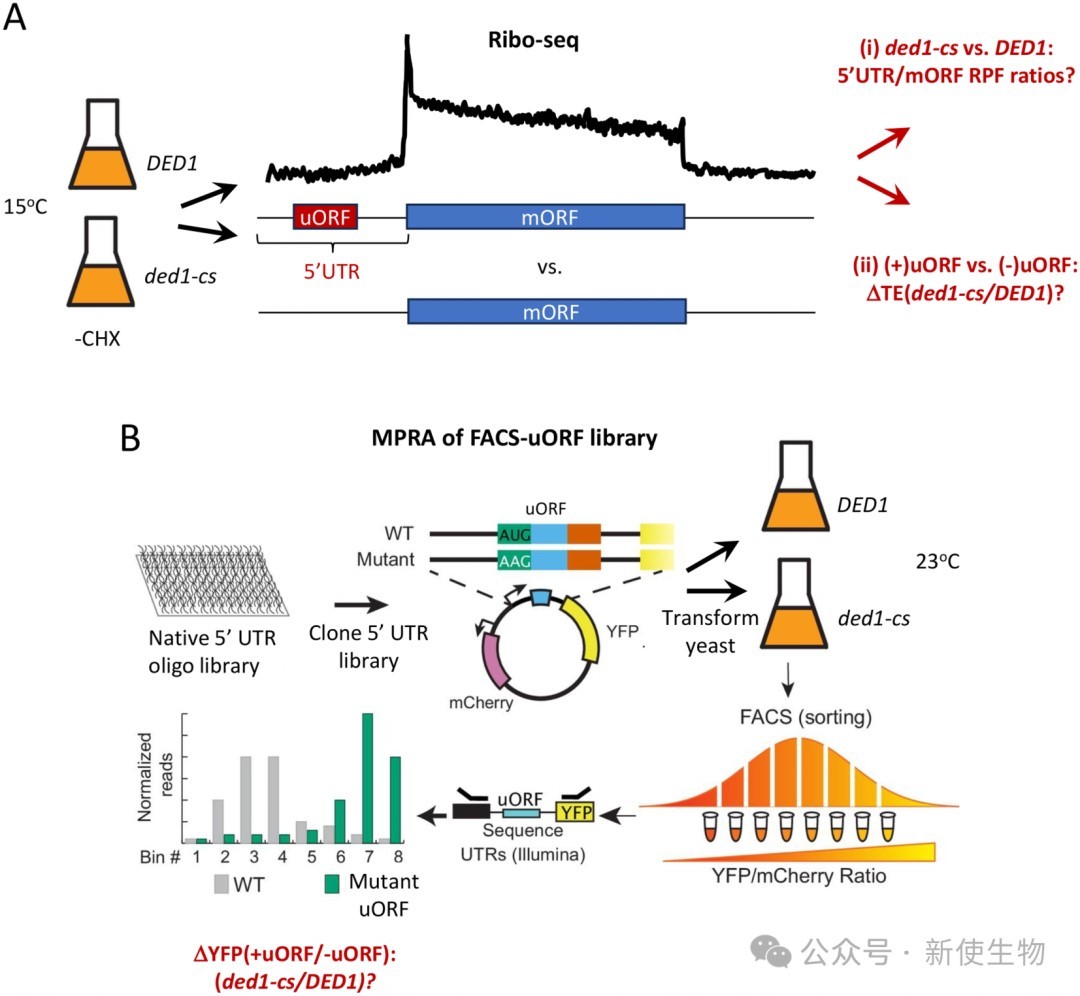
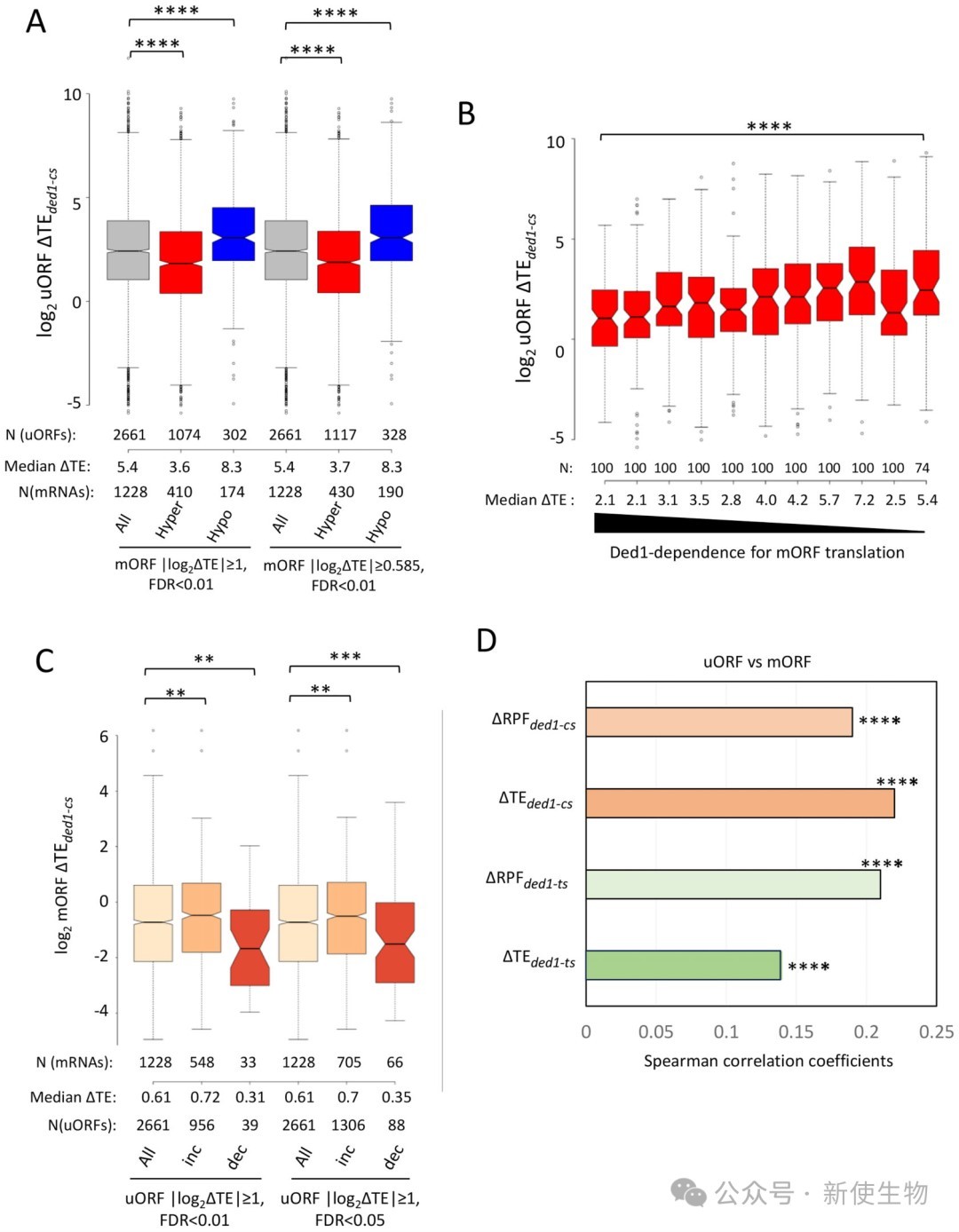
结果显示,在ded1突变体中,mORF翻译效率的降低并没有普遍伴随着其对应5' UTR中uORF翻译效率的增加,两者反而呈现正相关。
这表明Ded1突变导致的下游翻译抑制并非由上游uORF翻译的增强所致。
二、AUG起始的uORF不赋予下游mORF对Ded1的高度翻译依赖性
通过对不同标准鉴定的已翻译uORF进行分类分析,研究发现含有AUG起始uORF的mRNA在Ded1失活时并未表现出比无uORF的mRNA更显著的mORF翻译效率下降。
相反,只有含有NCC(非AUG)起始uORF的mRNA表现出一定的依赖性,但这种依赖性主要归因于这些mRNA本身具有更长且更具结构化的5' UTR,而非uORF本身。
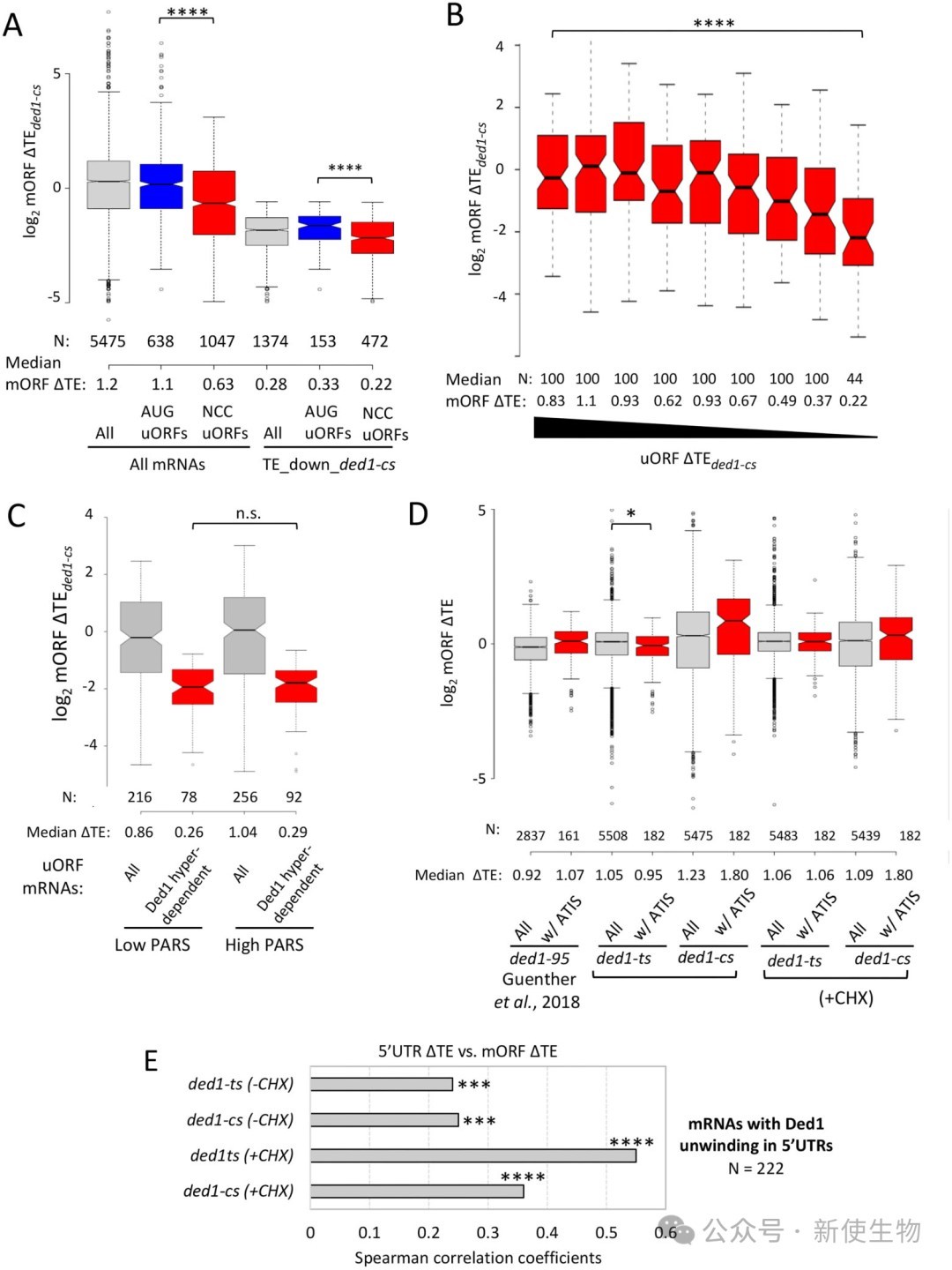
三、uORF下游的紧邻二级结构通常不介导对Ded1的依赖性
研究人员分析了uORF起始密码子下游不同区间内的RNA二级结构稳定性(PARS评分),发现uORF下游是否存在稳定的二级结构并不会加剧ded1突变对下游mORF翻译的抑制。
此外,对先前报道的Ded1解开的替代起始位点(ATIS)进行分析,发现这些位点在Ded1突变时也未表现出预期的翻译变化,表明下游结构辅助的uORF翻译抑制并非普遍机制。
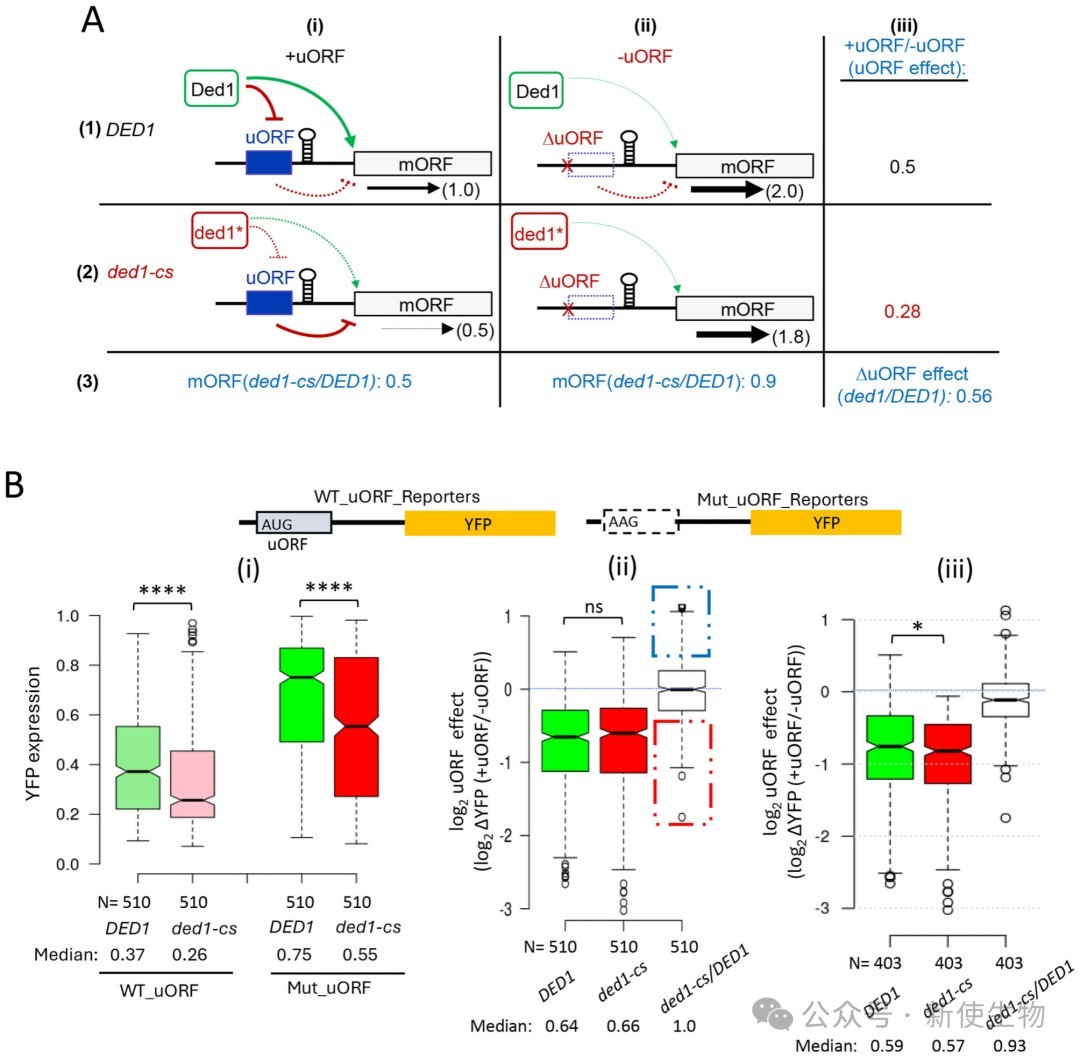
四、高通量报告基因分析证实uORF在ded1细胞中没有更强的抑制性
利用FACS-uORF高通量报告系统,研究人员在DED1和ded1-cs细胞中同时比较了数千个天然5' UTR及其uORF突变体的表达情况。
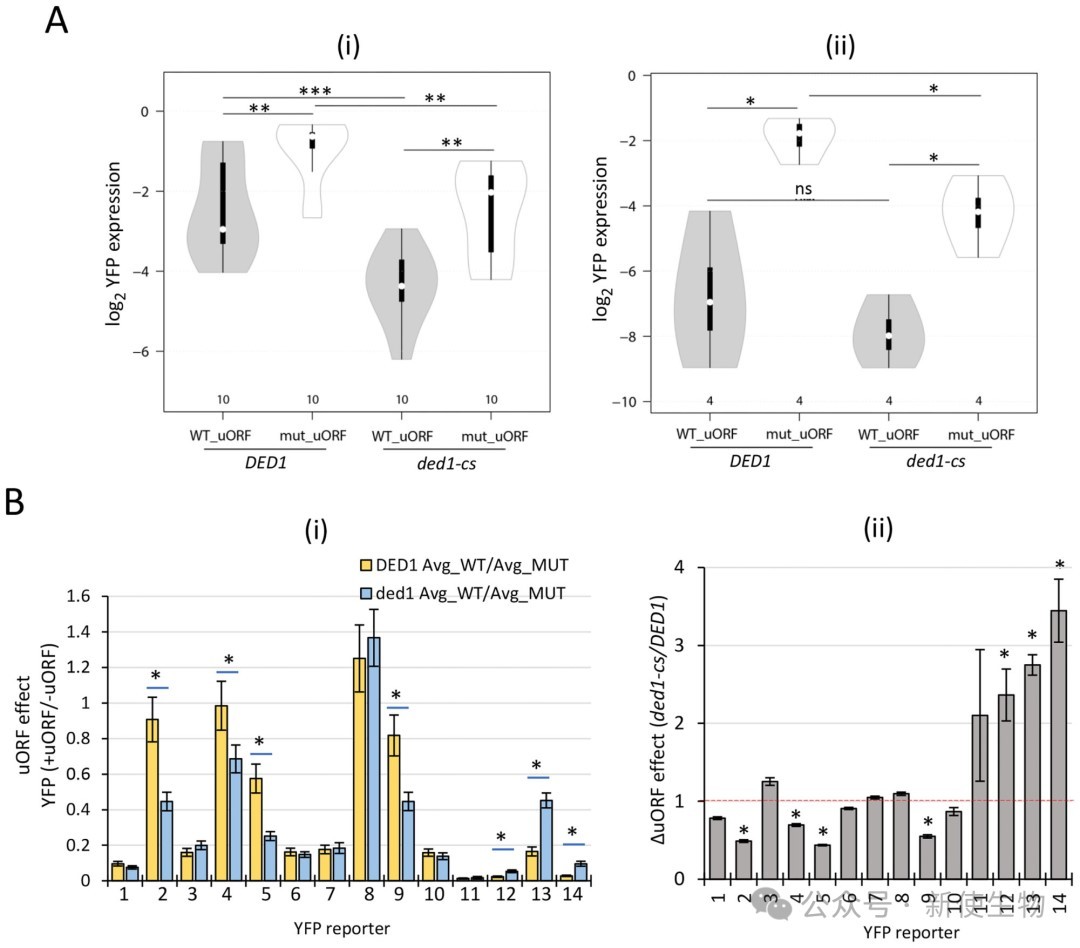
结果表明,绝大多数具有抑制性的uORF在Ded1功能缺陷时,其抑制强度并没有显著增加,说明Ded1并不普遍通过促进漏过扫描来克服uORF的抑制作用。
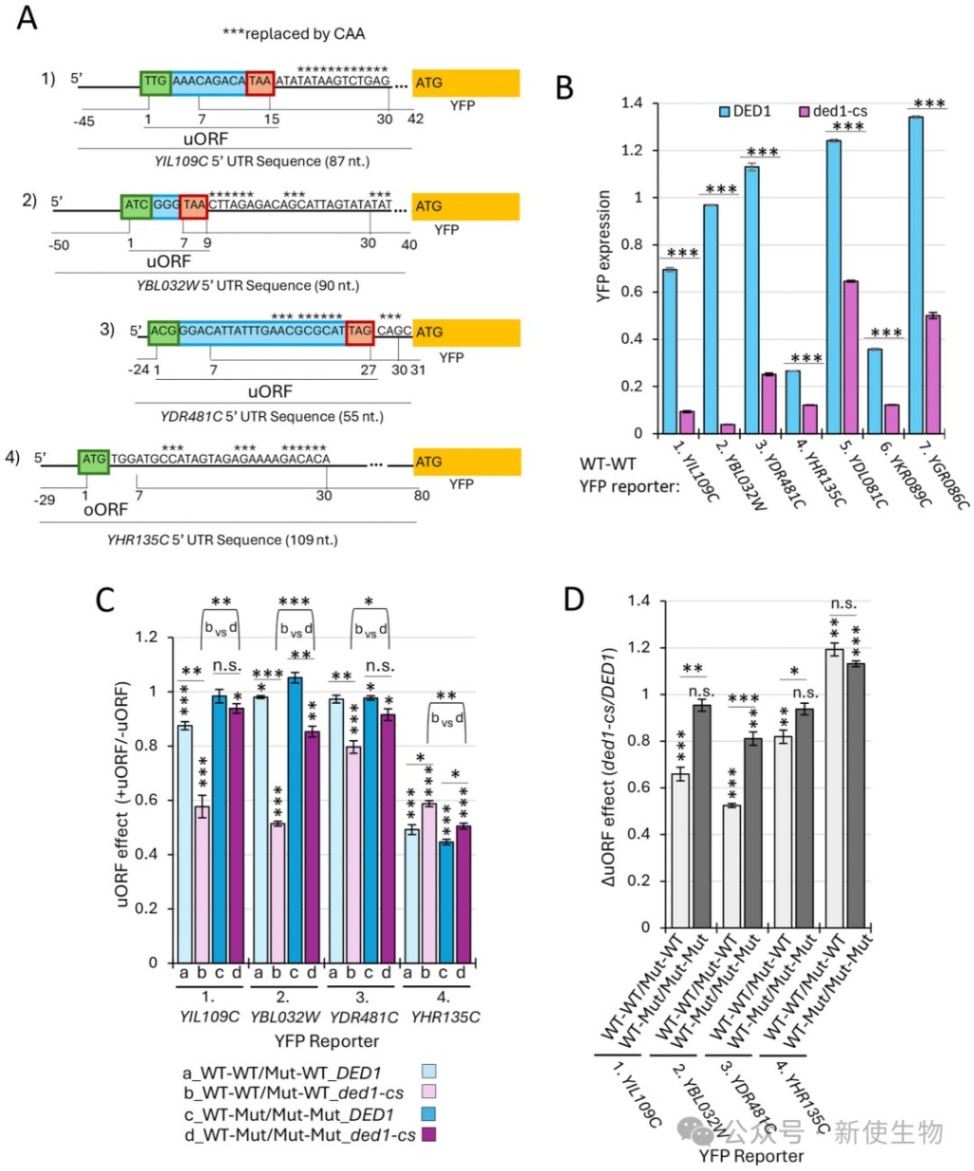
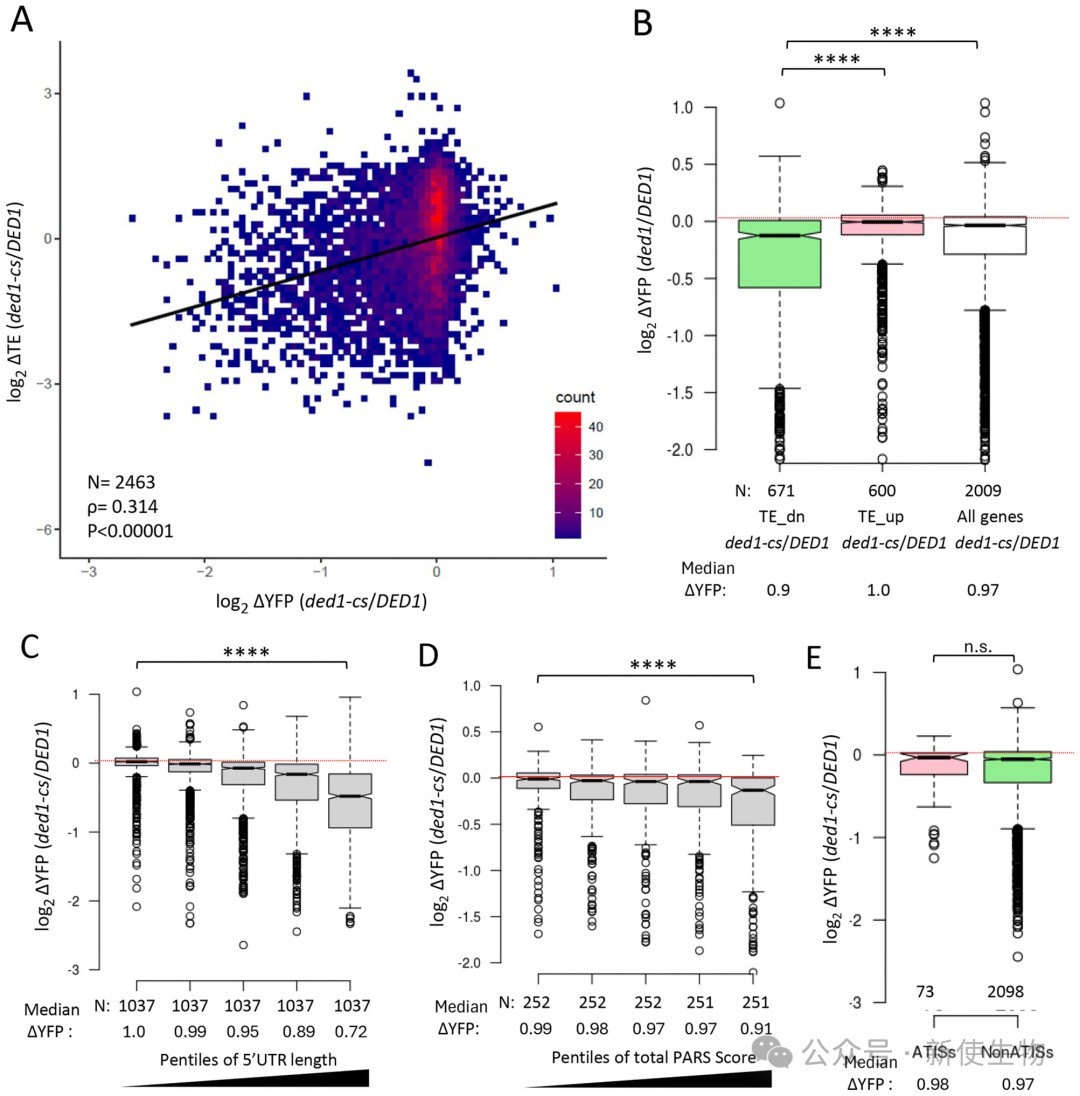
五、5' UTR的长度和结构复杂度是决定Ded1依赖性的主要因素
通过对数千个天然5' UTR报告基因的表达分析,研究发现随着5' UTR长度的增加、PARS结构的增强以及自由能的降低,报告基因对Ded1的依赖性显著增加。
即使在完全不含uAUG的报告基因中,这种由长度和结构决定的Ded1依赖性依然存在,进一步证实了解旋酶Ded1的主要功能是直接清除扫描通路上的物理障碍。
总结
本研究通过严谨的体内翻译谱分析和高通量报告系统,有力地反驳了Ded1主要通过抑制uORF翻译来促进主ORF翻译的假说。研究确立了Ded1作为通用解旋酶,其核心功能是直接解开5' UTR中的二级结构以促进43S前起始复合物的结合与扫描。这一结论为深入理解真核生物翻译起始的复杂调控网络提供了关键的分子证据。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看