

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
核糖体毒性应激反应(RSR)是细胞内的一条重要信号通路,由MAP3激酶ZAKα感知翻译过程中的异常并激活p38和JNK激酶。RSR的激活与细胞周期阻滞、程序性细胞死亡和炎症等多种应激结果直接相关。
多种环境和内源性应激源,如紫外线辐射、活性氧、蓖麻毒素和茴香霉素等,均可诱导RSR。近期研究表明,RSR通路在多种重要生物学现象中扮演关键角色,例如肿瘤发生、化疗敏感性和代谢调控。
尽管对RSR的研究兴趣日益浓厚,但其上游激酶ZAKα究竟如何感知翻译障碍,仍然是该领域悬而未决的核心科学问题。目前认为ZAKα可能识别碰撞的核糖体,也可能识别单个停滞的核糖体,但其精确的感知机制尚不明确。
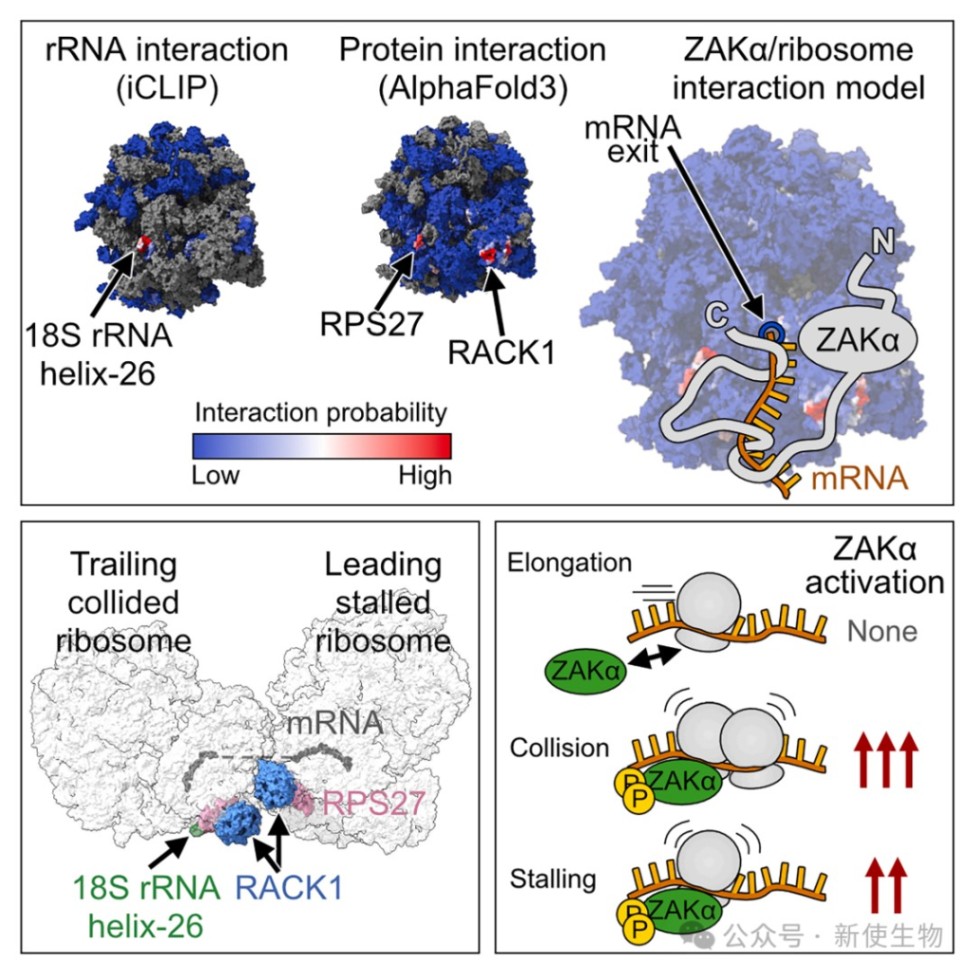
2026年6月13日,哥本哈根大学Simon Bekker-Jensen团队在Molecular Cell上发表了题为“Mechanism of RACK1-dependent ZAKα activation at stalled and collided ribosomes” 的研究论文。该研究揭示了核糖体毒性应激感受器ZAKα通过与核糖体上的RACK1、RPS27、18S rRNA以及mRNA出口通道处的mRNA形成一个复合结合界面,从而动态监控翻译进程。

文章索引
【标题】Mechanism of RACK1-dependent ZAKα activation at stalled and collided ribosomes
【发表期刊】Molecular Cell
【发表日期】2026年6月13日
【作者及团队】哥本哈根大学Simon Bekker-Jensen团队
【IF】16.6
研究结果
一、RACK1是ZAKα结合核糖体和激活的关键枢纽
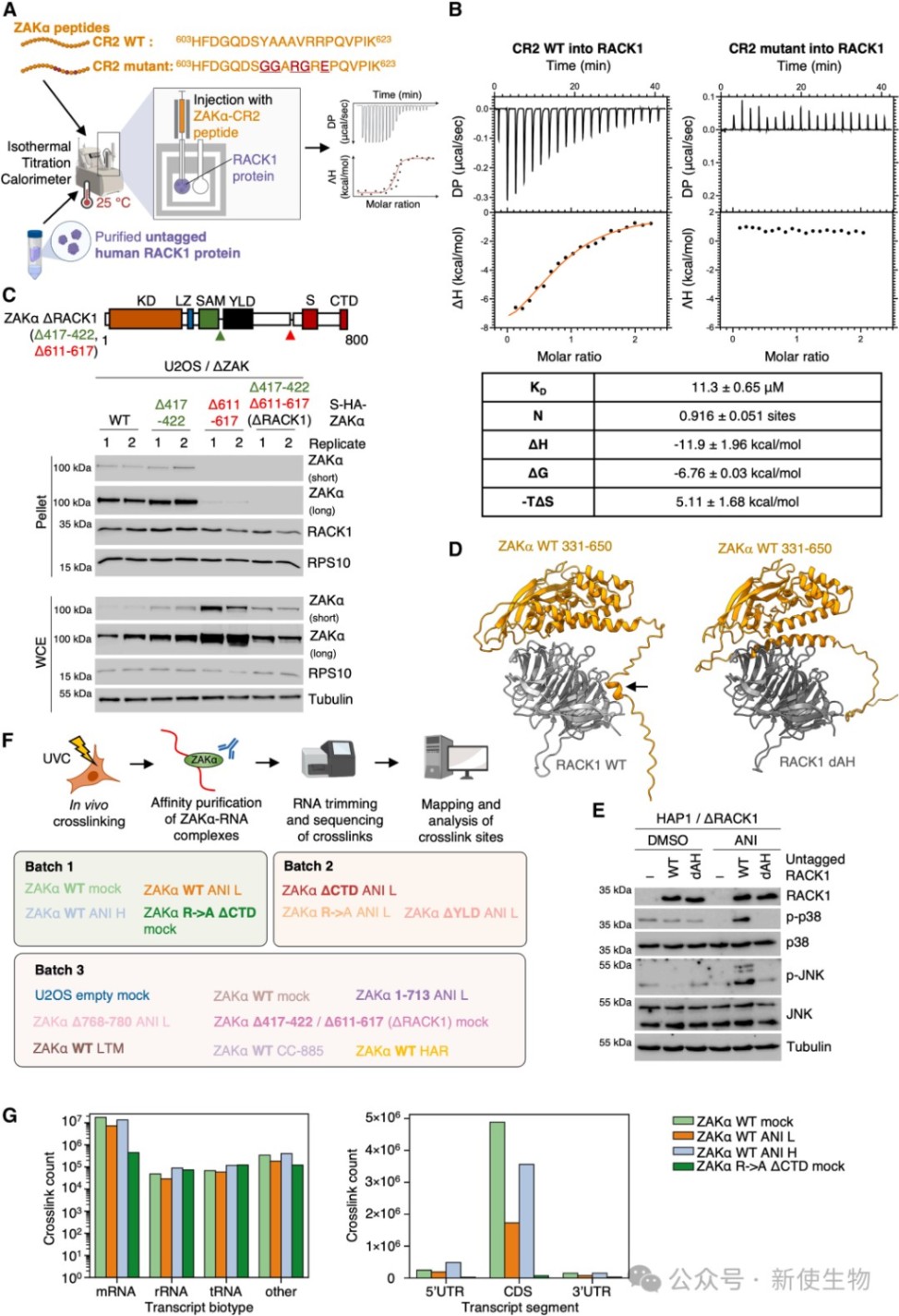
研究利用AlphaFold3预测并实验证实了ZAKα通过其C端的短线性相互作用基序(SLIMs)与核糖体蛋白RACK1和RPS27结合。
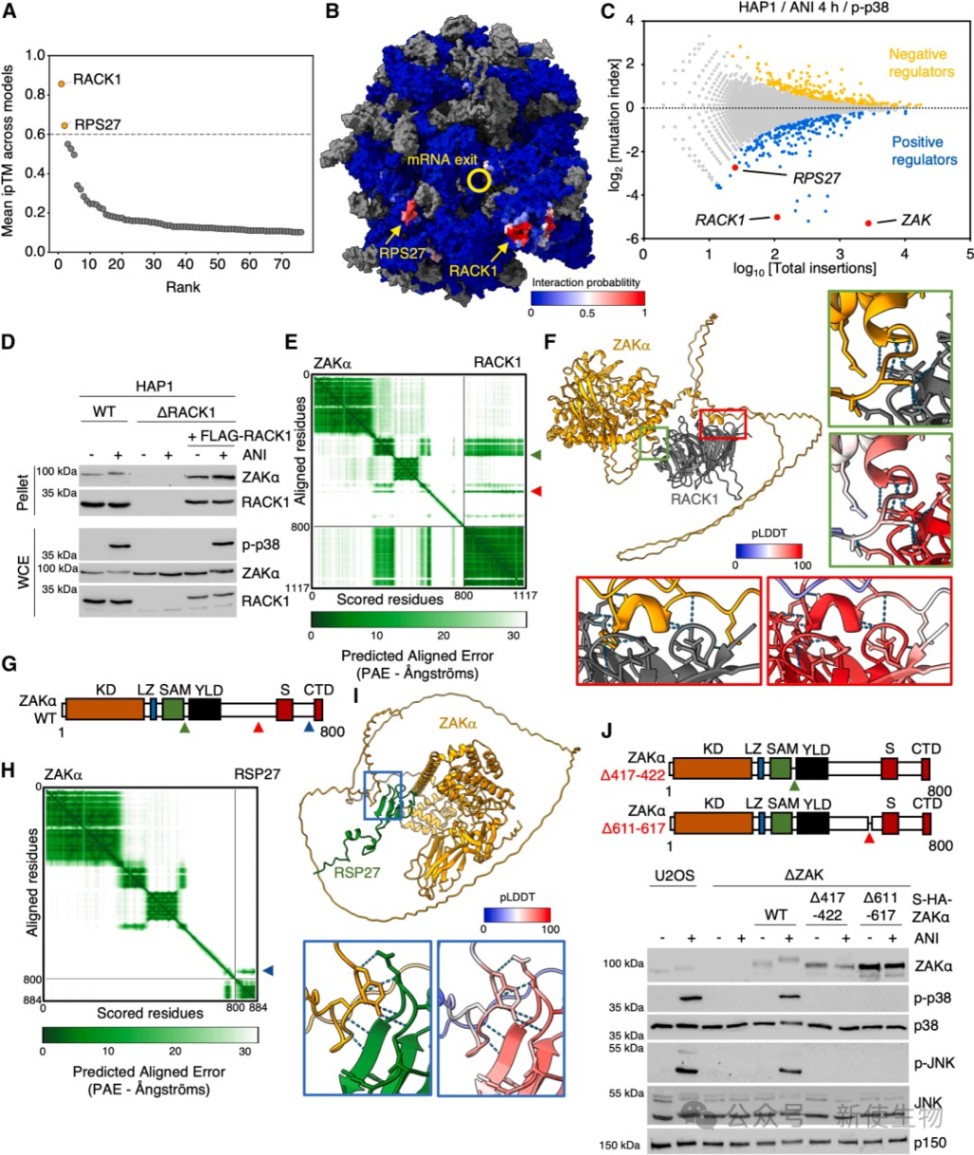
等温滴定量热法(ITC)和生化分析表明,缺失RACK1或突变ZAKα的特定基序会彻底破坏其与核糖体的结合及后续的p38/JNK激活。
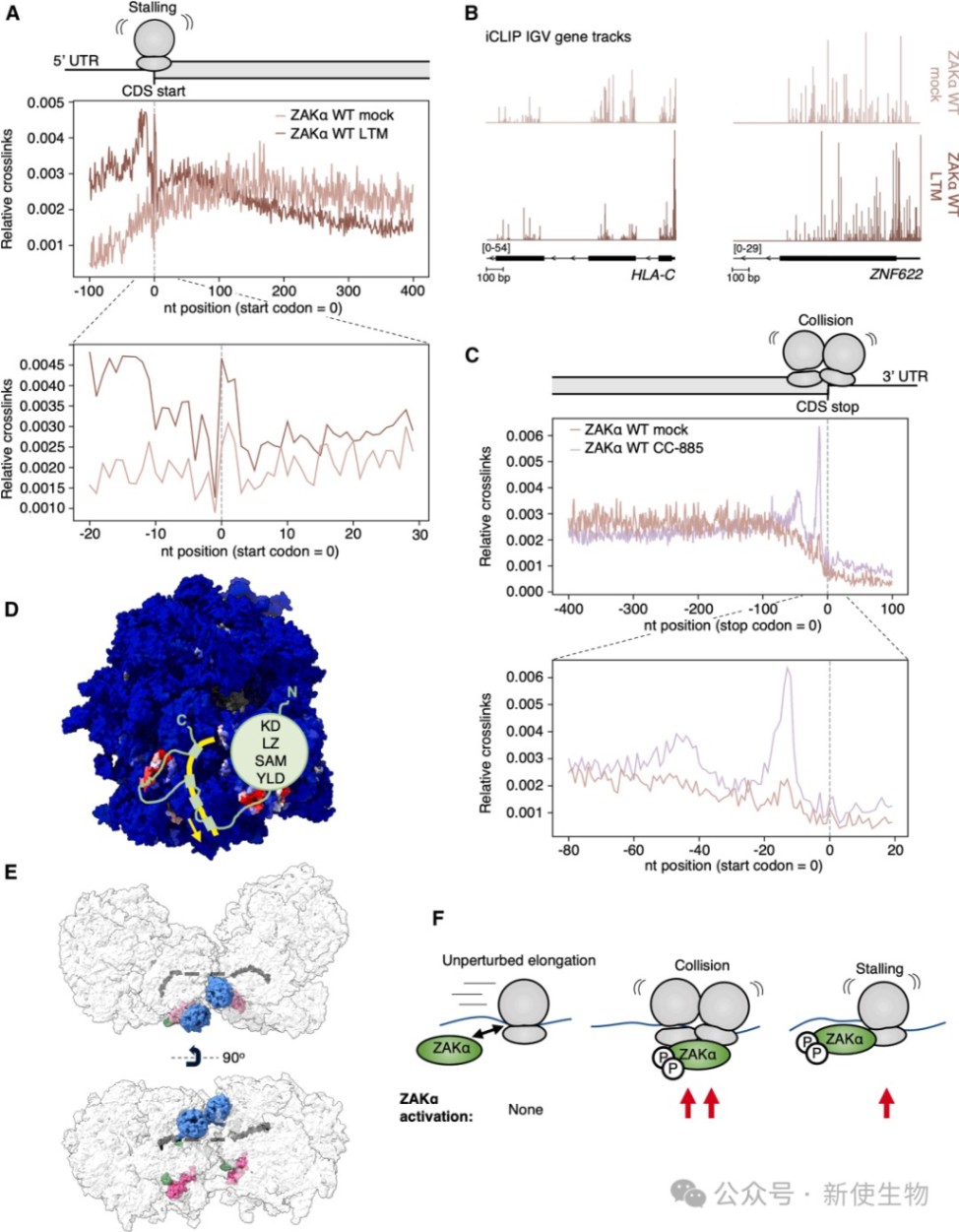
二、iCLIP揭示了ZAKα与18S rRNA helix-26的相互作用
通过iCLIP技术,研究人员在18S rRNA的helix-26区域发现了强烈的ZAKα交联信号。
这一位点在空间上邻近mRNA出口通道,证实了ZAKα在该区域的动态巡视模式,且无论在正常还是压力条件下,该相互作用均维持稳定。
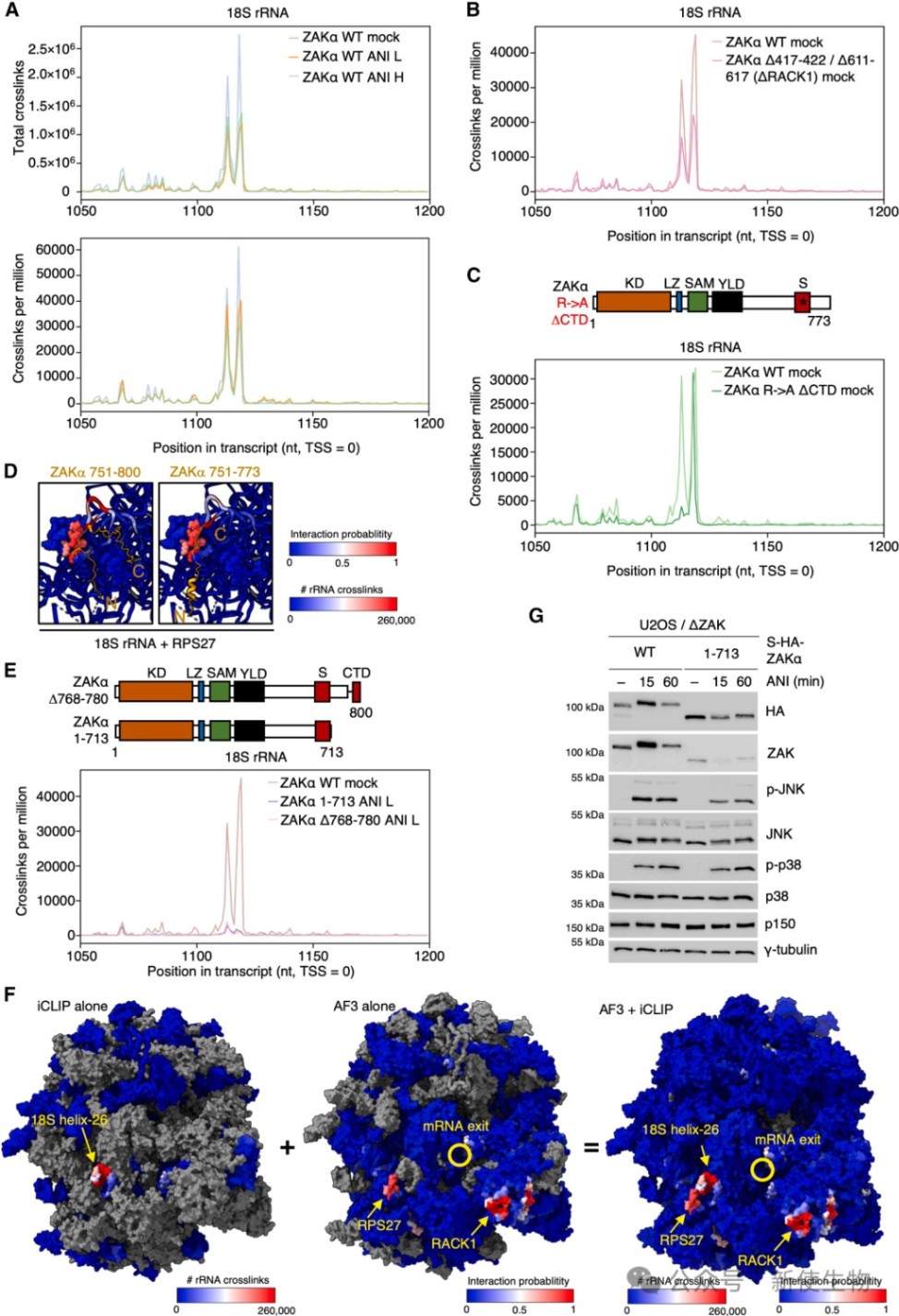
三、S和CTD结构域通过结合mRNA增强ZAKα与核糖体的亲和力
研究发现ZAKα的传感器(S)结构域和C端结构域(CTD)通过与穿过出口通道的mRNA直接接触来维持结合稳定性。
Disome-seq双核糖体印迹分析与iCLIP数据分析显示,ZAKα的mRNA交联图谱与核糖体在mRNA上的分布高度一致,证明其结合亲和力受mRNA状态的影响。
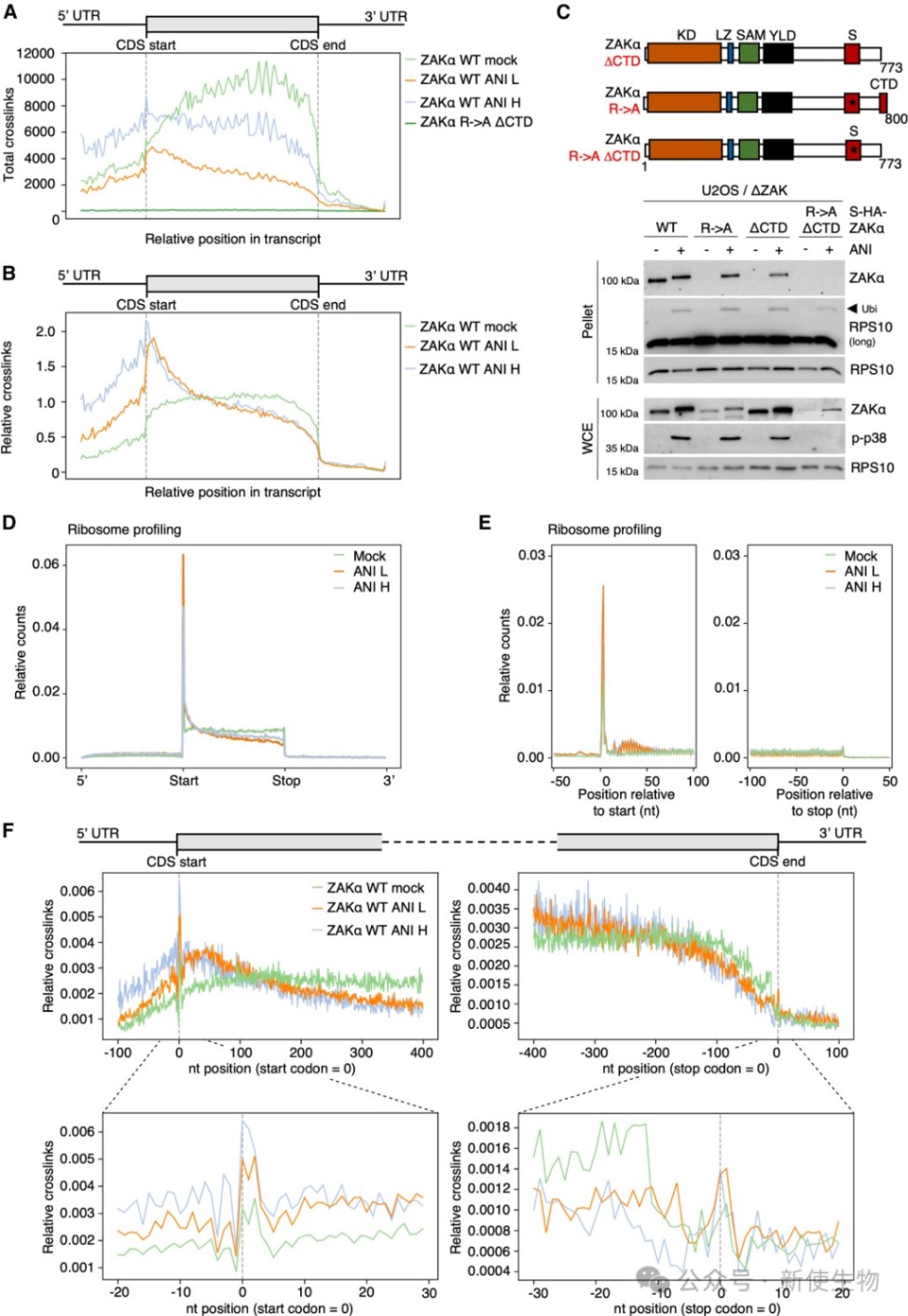
四、ZAKα可被单个停滞的核糖体以及碰撞核糖体激活
通过使用低剂量和高剂量的抗生素处理,Polysome profiling多聚核糖体分析证明在多种细胞系中,即使没有明显的核糖体碰撞,单个停滞的核糖体也足以触发ZAKα激活。
尽管碰撞核糖体能提供更强的信号强度,但感应机制本质上是基于核糖体合成过程的进程受阻。
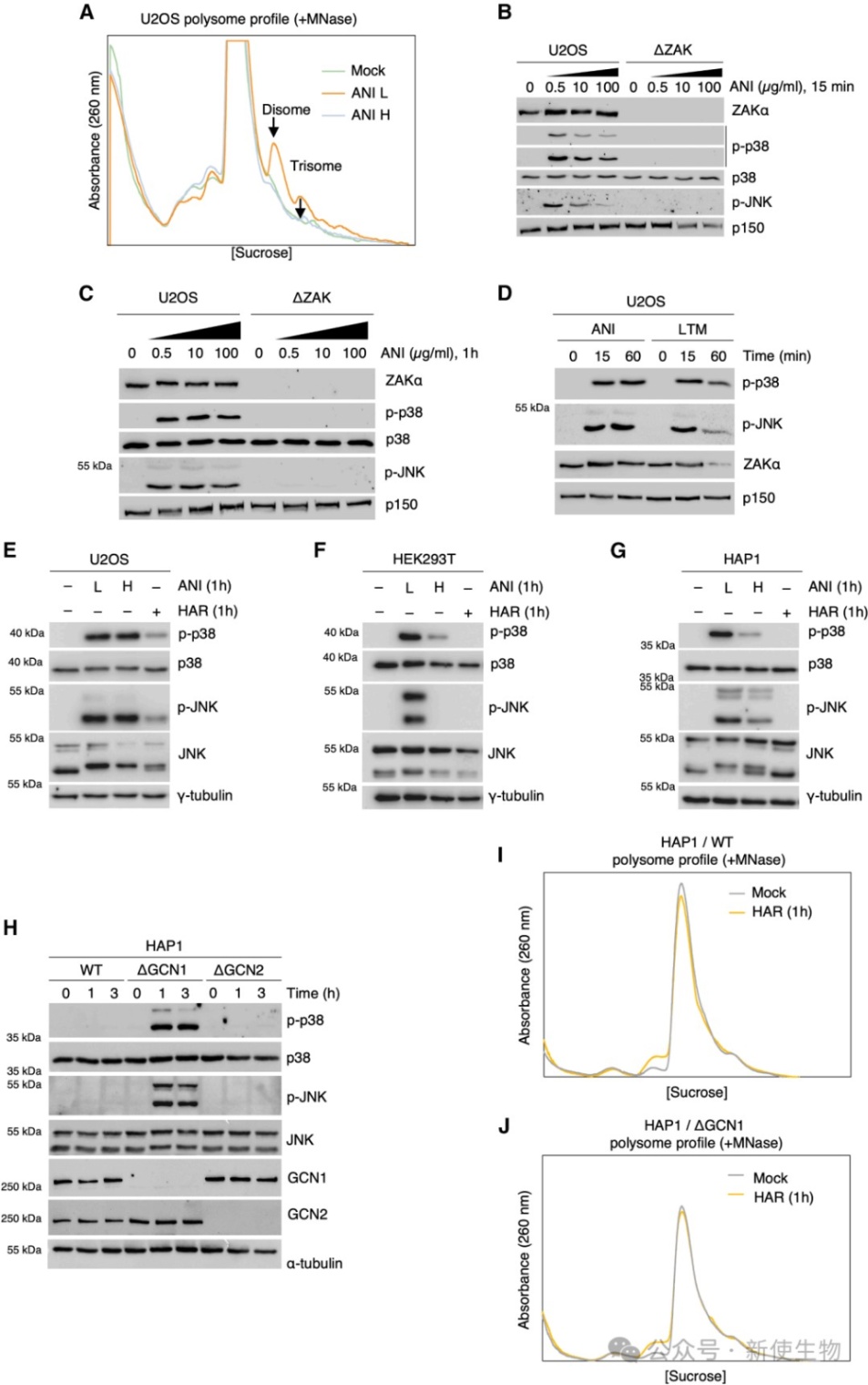
五、从核糖体上的解离受阻是ZAKα激活的前提条件
在正常翻译中,核糖体的持续移动会不断将ZAKα从结合位点推开,使其保持单体非激活状态。
当发生核糖体停滞或碰撞时,mRNA的静止导致ZAKα在核糖体上驻留时间延长,从而为后续的二聚化和激活提供了窗口。
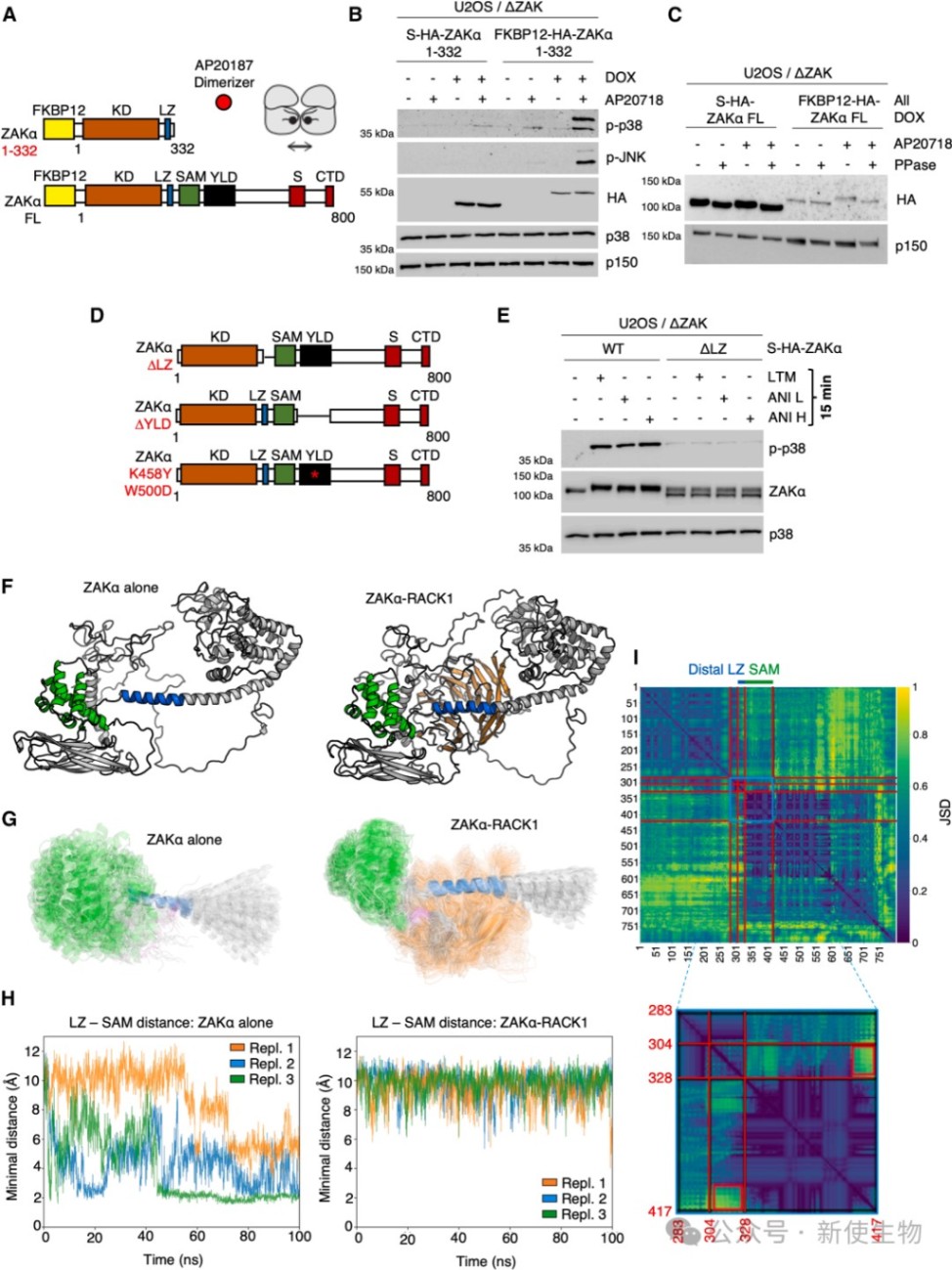
六、核糖体模板诱导的ZAKα二聚化和构象变化驱动激酶激活
分子动力学模拟显示,ZAKα结合RACK1后会隔离其抑制性的SAM结构域,从而释放亮氨酸拉链(LZ)结构域以促进二聚化。
这种由核糖体作为模板诱导的近距离接触促使激酶结构域发生反式自磷酸化,最终启动下游应激信号传导。
总结
本研究详细刻画了ZAKα在核糖体mRNA出口通道处的分子图谱,证明其通过协同结合核糖体蛋白、rRNA和mRNA来监控翻译进程。这种多位点结合模式使得ZAKα能够灵敏地捕捉到mRNA的停滞信号,通过在受损核糖体上的稳定驻留和二聚化完成信号转导,从而统一了关于核糖体停滞与碰撞触发应激反应的理论模型。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看