

 首页
首页 关于我们
关于我们 产品中心
产品中心 技术服务
技术服务 技术中心
技术中心 联系我们
联系我们
导读
过去十年中,原核生物与真核生物的抗病毒免疫机制展现出惊人的相似性,揭示了两者之间深层的进化联系。这些保守的免疫策略包括由cGAS类酶产生的环状核苷酸信使以激活下游效应器,以及形成孔道复合物诱导细胞死亡。
此外,模式识别受体和链终止核苷酸的产生也被证明是跨物种的防御手段。哺乳动物的Schlafen(SLFN)核酸酶通过干扰素诱导表达,能够切割宿主tRNA和rRNA以抑制病毒感染。
SLFN11是其中研究最深入的成员,它通过在密码子偏好性位点切割tRNA来限制HIV复制。然而,尽管Schlafen结构域广泛存在于细菌和古菌中,其在哺乳动物之外的生物学功能和作用机制长期以来尚不明确。
2026年7月2日,哈佛马省总医院Jonathan Strecker团队在 Nature Communications 上发表了题为“Prokaryotic Schlafen proteins cleave tRNAs during type III CRISPR immunity”的研究论文。该研究鉴定并表征了一类新型CRISPR相关Schlafen蛋白质(Cash),发现其作为III型CRISPR系统的效应因子,受cA₄信号分子激活并主要在T-loop区切割宿主tRNA。

文章索引
【标题】Prokaryotic Schlafen proteins cleave tRNAs during type III CRISPR immunity
【发表期刊】Nature Communications
【发表日期】2026年7月2日
【作者及团队】哈佛马省总医院Jonathan Strecker 团队
【IF】15.7
研究结果
一、Cash蛋白质受III型CRISPR系统激活并诱导细胞毒性
研究者在III型CRISPR位点中发现了由Schlafen结构域和类Rossmann传感器结构域Csx15融合而成的Cash蛋白。
实验证明Cash受Cas10产生的环腺苷酸信号激活,通过切割tRNA(主要是T-loop区)诱导细胞毒性,从而通过流产感染机制阻止病毒扩散。
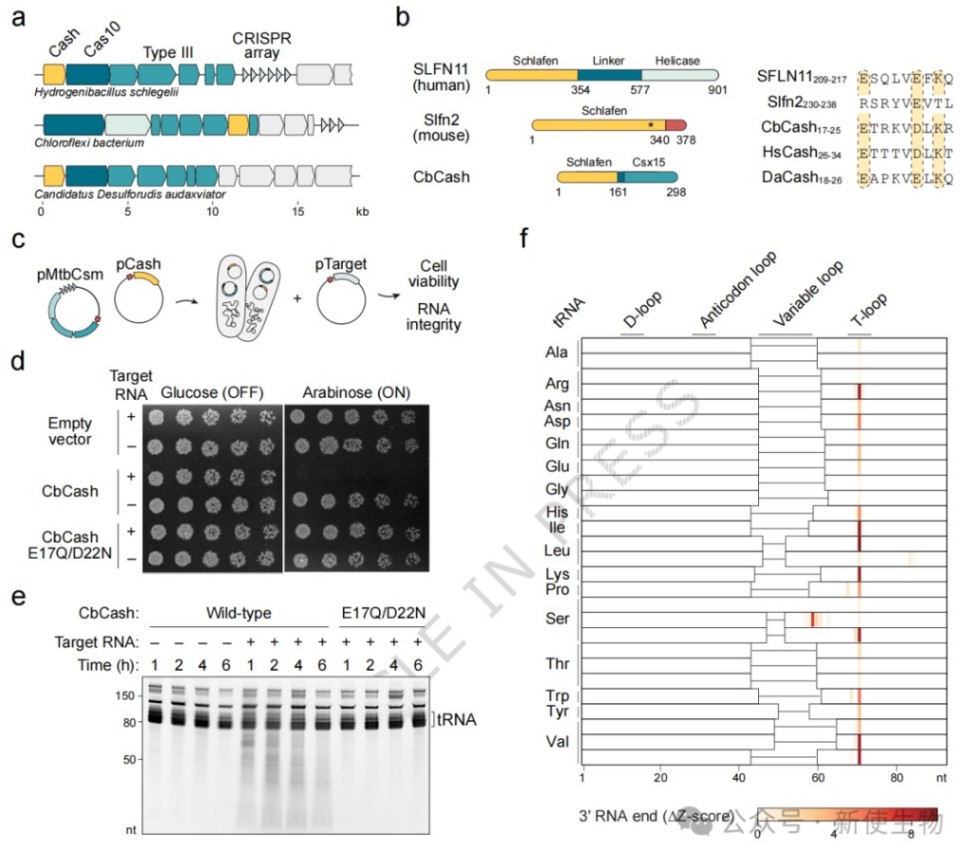
二、CbCash是一种受cA₄触发的tRNA内切核酸酶
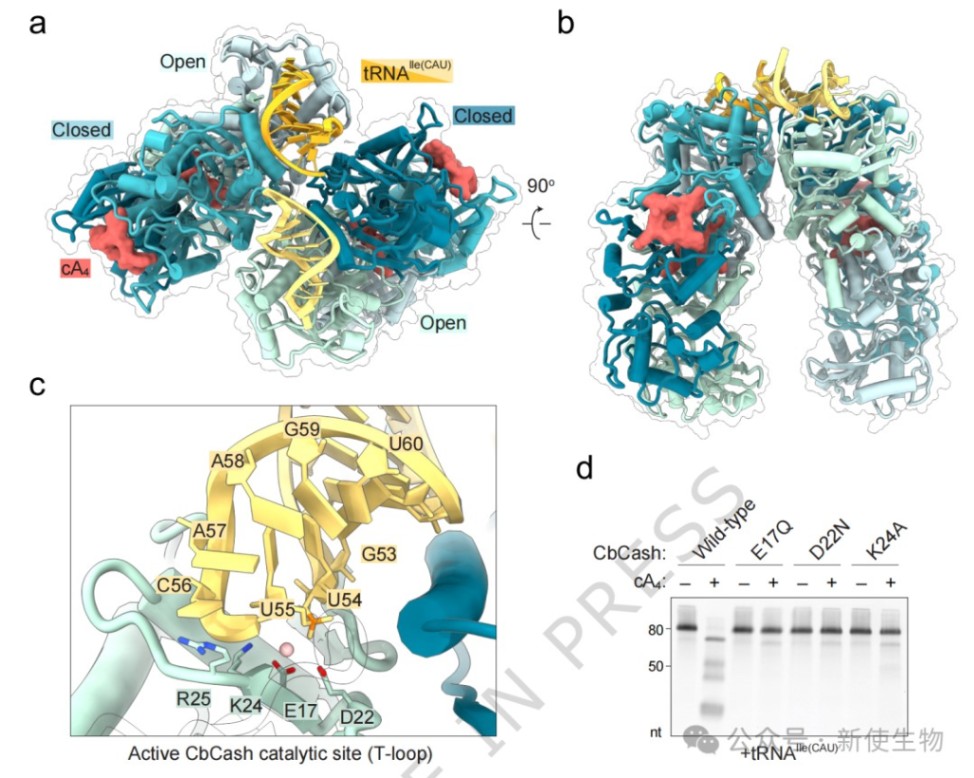
体外生化实验表明,CbCash的生化活性严格依赖于环四腺苷酸(cA₄)及其催化位点残基。
tRNA-seq显示其广泛切割宿主tRNA,识别tRNA环中保守的U-turn结构和UUC基序,并将其水解为特定的小分子片段。
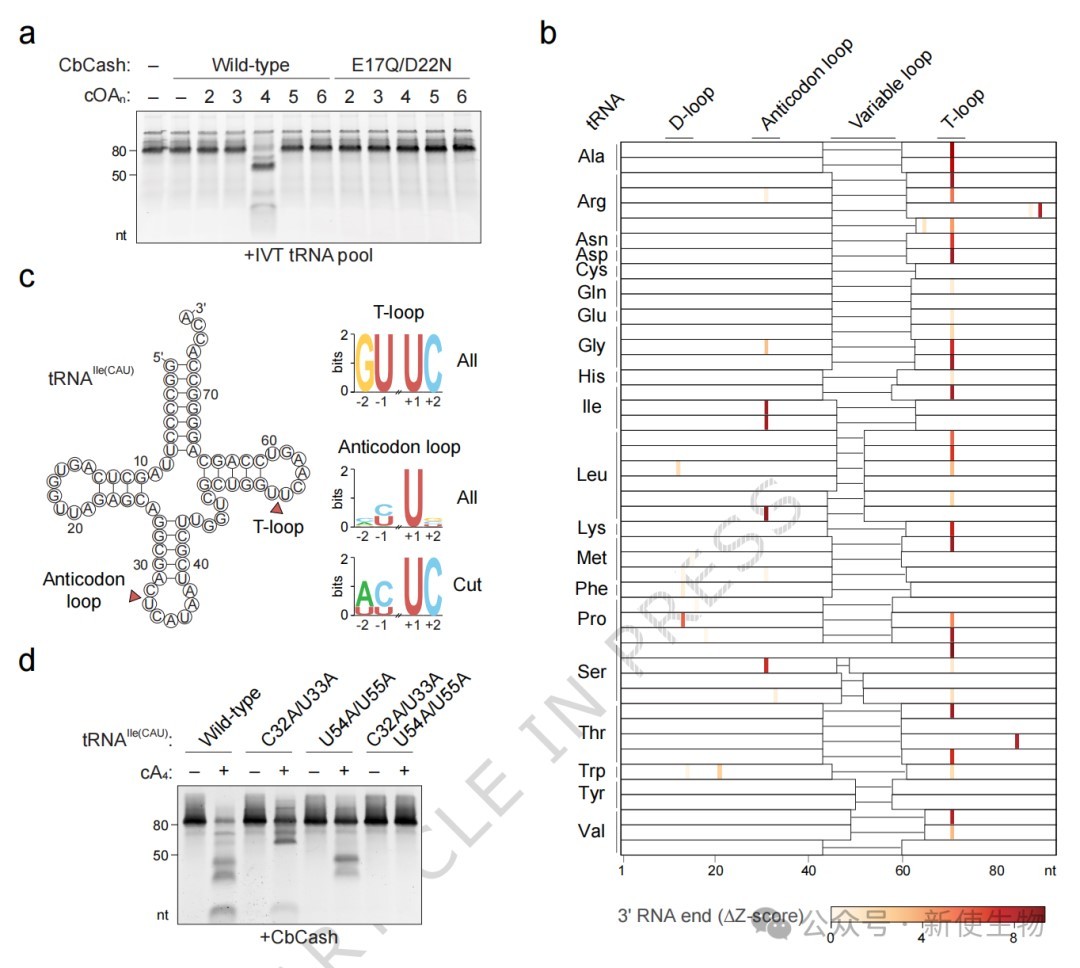
三、CbCash在静息状态下形成不活跃的十二聚体
利用冷冻电镜技术,研究者解析了CbCash在apo状态下的2.8Å结构,发现其由六个二聚体组装成具有D2对称性的十二聚体。
这种复杂的架构通过高度稳定的盐桥网络固定,将Schlafen结构域锁定在封闭状态,从而抑制其背景催化活性。
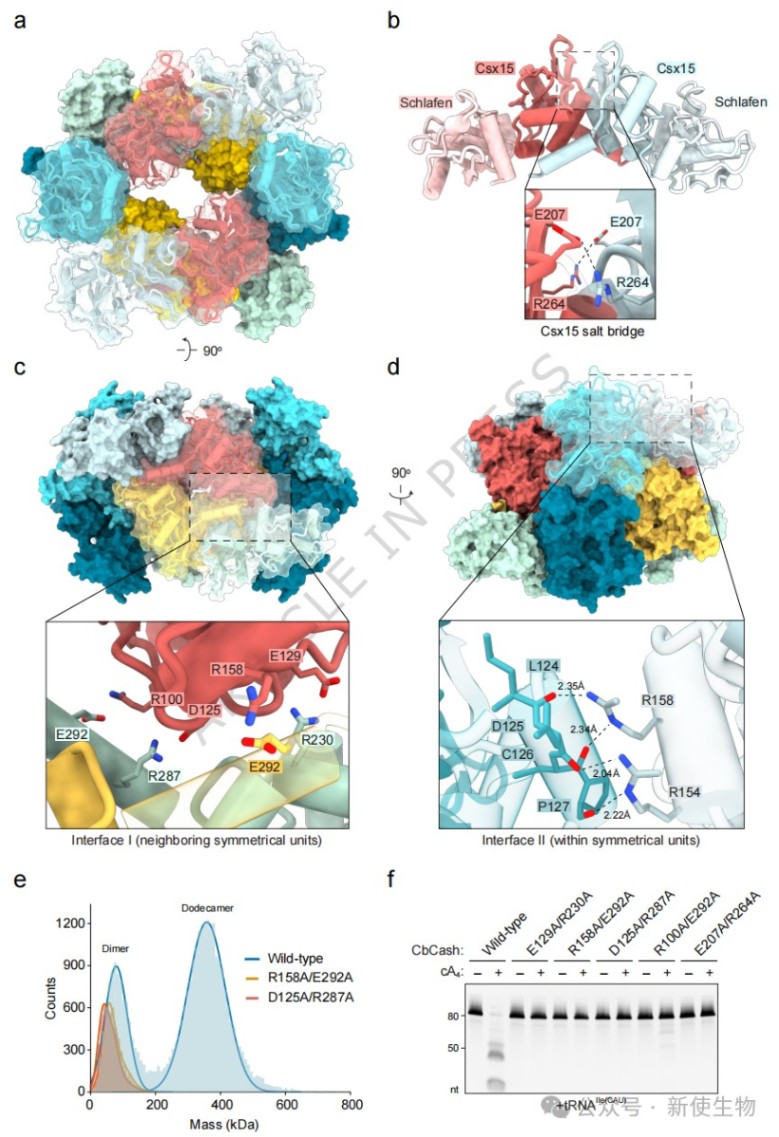
四、cA₄诱导CbCash发生丝状化转化
在加入cA₄后,CbCash十二聚体迅速解离并重新聚合成长丝状结构,冷冻电镜显示该长丝由平行的Schlafen二聚体重复单元构成。
cA₄结合在Csx15界面处诱导了显著的构象改变,使Schlafen结构域由“封闭”转为“开放”状态,从而暴露活性催化中心。
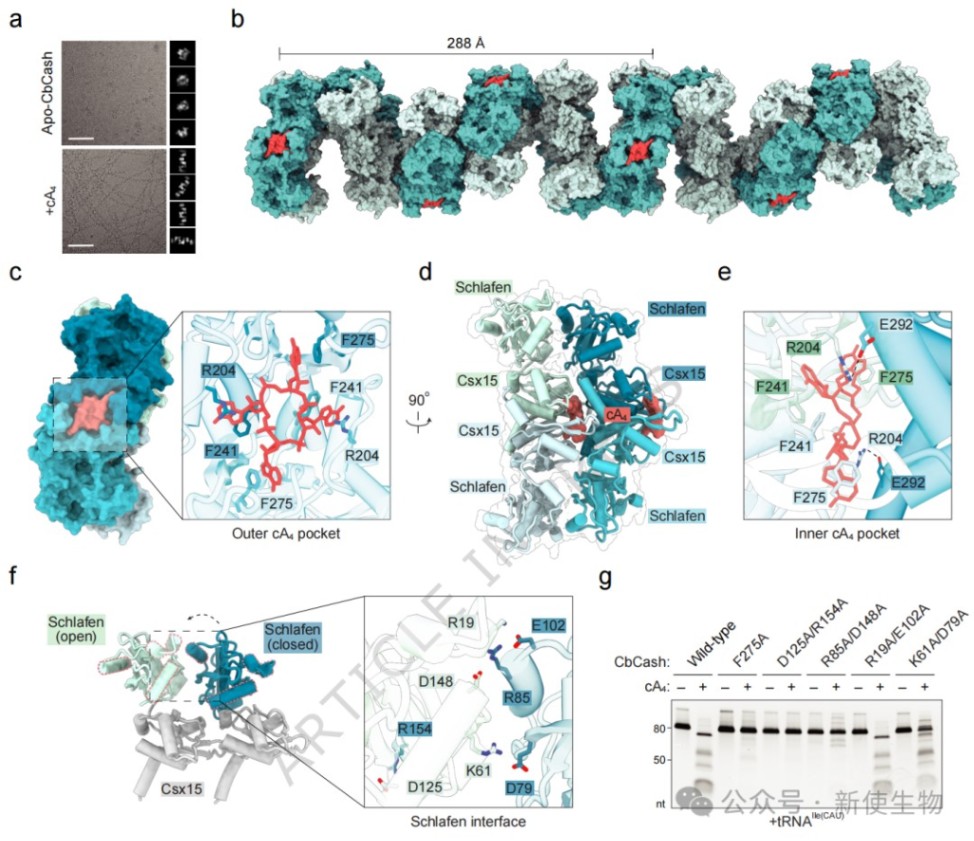
五、CbCash识别与切割tRNA的分子基础
通过解析tRNA结合状态的CbCash长丝结构,研究者观察到tRNA的特定环状结构被精准定位于开放的Schlafen口袋内。
活性中心通过E17-D22-K24催化三元组和镁离子,定位tRNA回折处的磷酸二酯键并执行剪切。
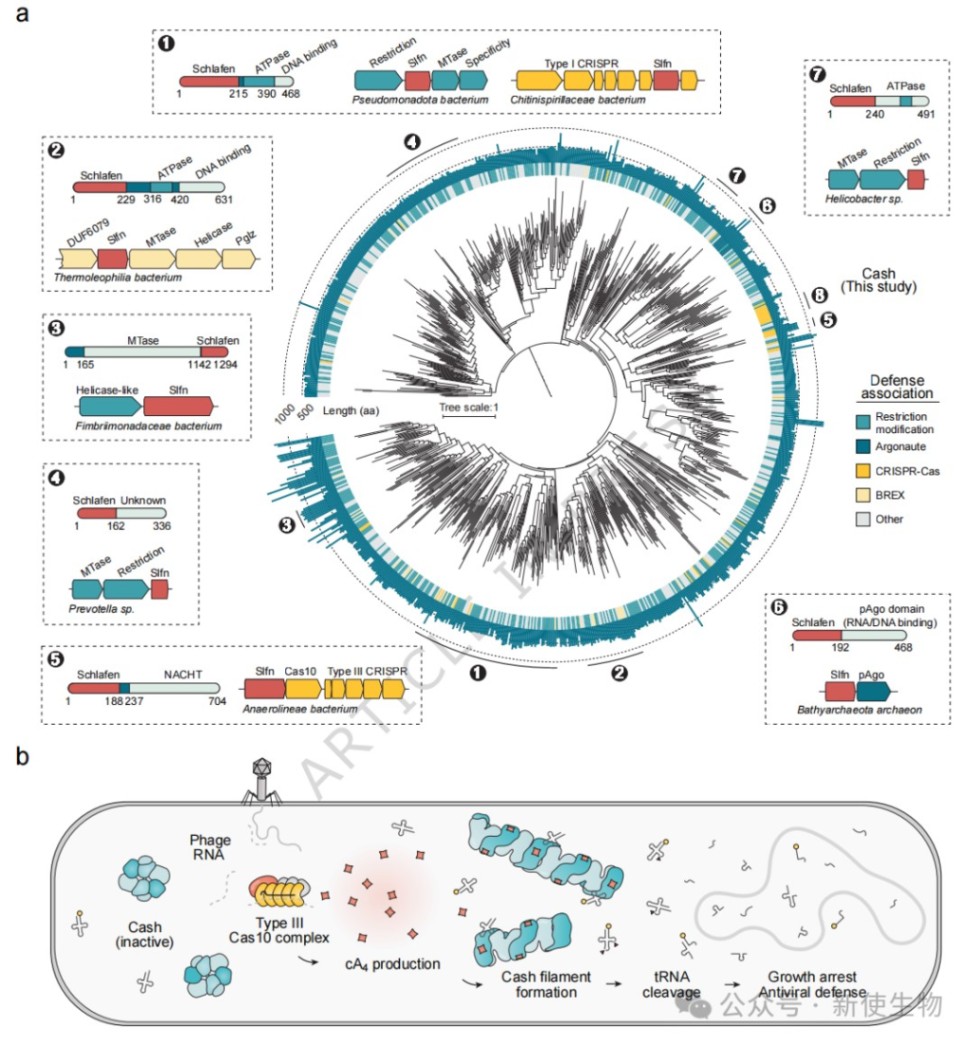
六、Schlafen蛋白质在原核生物防御系统中广泛分布
生物信息学分析揭示了多种与原核防御系统相关的Schlafen同源物,它们与Argonaute、限制修饰系统或III型CRISPR系统紧密关联。
这些蛋白质具有多样的传感器结构域,表明Schlafen核酸酶在原核免疫中具有多种多样的触发机制。
总结
本研究系统地表征了一类原核Schlafen蛋白(Cash)的结构与功能,揭示了它作为III型CRISPR免疫系统下游效应子的新角色,其通过cA₄信号激活、组装成纤维丝并切割tRNA来抵抗病毒感染。这项工作不仅扩展了我们对CRISPR免疫机制的理解,也证明了Schlafen核酸酶介导的tRNA消耗是贯穿原核生物与真核生物的一种古老且保守的抗病毒策略。
| 新使生物专业翻译组一站式服务平台 |
| 产品名称 |

点击图片查看
点击图片查看